https://doi.org/10.22319/rmcp.v16i4.6744
Article
Chemical composition and adaptation of tropical grass Leersia hexandra Sw. exposed to crude oil soil
José Alberto Orocio-Carrillo a*
María del Carmen Rivera-Cruz a
Said Cadena-Villegas a
Consuelo del Carmen Bautista-Muñoz a
Antonio Juárez-Maldonado b
Karla Chávez-Álvarez a
a Colegio de Postgraduados. Campus Tabasco. Programa de Doctorado en Ciencias Agrícolas en el Trópico, Laboratorio de Microbiología Agrícola Ambiental. Periférico Carlos A. Molina, Km 3.5. 86500, H. Cárdenas, Tabasco, México.
b Universidad Autónoma Agraria Antonio Narro. Departamento de Botánica. Coahuila, México.
* Corresponding author: orocio.alberto@colpos.mx
Abstract:
The purpose was to evaluate the chemical composition and adaptation potential of L. hexandra under crude oil stress conditions, through the rhizobacterial population, crude protein accumulation, neutral detergent fiber, acid detergent fiber, and lignin in foliage of young plants emerging from the main tillers of the plant at different growth ages (d 180 and 360), as well as the production of young plants in the tillering stage, and the aerial and root dry matter. The results showed that crude oil concentrations in the soil significantly affected the population of Azotobacter spp. (0.361*); however, those of Azospirillum spp. and Pseudomonas spp. were inhibited, while both populations increased with time extension (0.778*, 0.767*). Likewise, the synthesis of crude protein (0.551**) and lignin in the foliage (0.354*) and the production of young plants in the tillering stage (0.465**), as well as of root dry matter (0.362*) were increased, indicating a strategy of L. hexandra to survive and adapt to soil contamination by crude oil. Nevertheless, the chemical composition was affected by the age of the grass, in which the percentage of neutral detergent fiber (0.832**), acid detergent fiber (0.741**), and lignin (0.661**) increased, while that of crude protein decreased (-0.497**).
Keywords: Rhizobacteria, Crude Protein, Lignin, Neutral Detergent Fiber.
Received: 07/07/2024
Accepted: 07/10/2024
Introduction
Soils contaminated with crude oil have generated a toxic environment for plants and microbial activity in the rhizosphere of various plant species(1,2) due to petroleum’s toxic, mutagenic, and carcinogenic nature(3). Soil contamination with crude oil affects fertility in a negative way because crude oil decreases the availability of nutrients (nitrogen, phosphorus, and potassium), pH, and moisture content, and increases the electrical conductivity and organic carbon content of the soil(4) directly affecting plant growth and development(5). In addition, the oil also forms a hydrophobic surface around the roots, which limits the absorption of water and nutrients from the soil(6). In oiled soil, Leersia hexandra grows and produces forage(7); it is a perennial grass that inhabits humid areas in tropical and subtropical regions; it propagates vegetatively by rhizomes, invading wetlands by the abundant accumulation of its foliage and roots on the soil and water(8) and is highly consumed by cattle in both dry and rainy seasons(9).
The rhizosphere is the soil-root interface, the volume of soil influenced by root activity(10). Tropical grasses are characterized by an extended rhizosphere system due to the abundant fibrous root system that characterizes them, and have demonstrated adaptation to the stress conditions established in soils contaminated with crude oil(11). Tropical grasses that have naturally evolved under stress conditions due to the accumulation of biogenic and petroleum carbon in the soil are able to mitigate stress by changing the chemical conditions in the rhizosphere and the proliferation of specific heterotrophic microorganisms(12,13). The rhizobacteria are free-living heterotrophs associated with the plant root(14); they stimulate plant growth, provide plant tolerance to petroleum diesel stress(15), and are involved in the rhizodegradation processes of alkanes, cycloalkanes, and polycyclic aromatic hydrocarbons(16). Azotobacter, Azospirillum, and Pseudomonas bacterial genera tolerate crude oil-induced stress(17,18). Plant growth-promoting rhizobacteria that colonize the rhizosphere mineralize intermediate metabolites from the decomposition of organic carbon in petroleum(19), and the exoenzymes released by them reduce and oxidize nitrogen, phosphorus, and sulfate, but also fix nitrogen biologically(20). The adaptation of grasses to crude oil soil suggests that it has a rhizosphere with roots that assimilate NO3, H2PO4, and SO4, and are used in an essential way in the primary metabolism inside the plant(21). Grasses adapted to crude oil exposure are likely to respond in a similar manner to other plant species such as Banksia seminuda Rie., Hakea prostrata R. Br.(22), and Secale cereal L.(23) where abiotic stress modifies the development and expression of genes involved in cell synthesis in the roots and foliage(24). The adaptation of plastids to stress by abiotic factors is based on the positive response of growth, production, and synthesis of secondary metabolites. In this regard, Orocio-Carrillo et al(13) report that the root and leaf protein content of L. hexandra has a hormetic response to doses of total petroleum hydrocarbons in the soil. Correa and Maranho(25) indicated that stem length, root biomass, and stomatal density increase in Echinochloa polystachya (Kunth) Hitchc. exposed to oil-containing soil; Habermann et al(26) indicate that water deficit stress and soil heating induced an increase in fiber and lignin content and reduced leaf protein in Panicum maximum Jacq; other authors(27,28) report that high temperatures lead to high detergent fiber and lignin content in the cell wall, and decreased protein synthesis in forage grasses. The objective of this study was to determine the rhizobacterial population, crude protein accumulation, neutral detergent fiber, acid detergent fiber, and lignin in the foliage, as well as the production of young L. hexandra plants in the tillering stage and in their aerial and root dry matter, to obtain a grass adapted to soils contaminated with crude oil for the Mexican humid tropics.
Material and methods
Soil and rhizome collection
Uncontaminated soil (Gleysol) was collected from the surface layer (0-30 cm) located in Ejido Blasillo 4th Section, Huimanguillo, Tabasco, Mexico (18° 05' 08.4” N and 93° 56' 50” W). The soil was dried under shade, ground, and sieved (5 mm mesh). The physical and chemical characteristics of the soil are shown in Table 1. L. hexandra rhizomes were collected from a wetland affected by a chronic oil spill, located two kilometers southwest of the “La Venta” Gas Processing Complex. The cultivation of L. hexandra seedlings was similar to the procedure used by Orocio-Carrillo et al(13).
Table 1: Physical and chemical characteristics of soil and oil
OM= organic matter; TN= total nitrogen; EC= electrical conductivity; CEC= cation exchange capacity. aHydrocarbon fractions using the Soxhlet and gravimetric methods.
Soil contamination and experimental design
32° API crude oil (CO) was obtained from the Ogarrio Field, Battery 2, in Huimanguillo, Tabasco, Mexico. The experiment was carried out with a completely randomized design and a 4x2 factorial arrangement: four concentrations of CO [0 (control), 30, 60, and 90 g kg-1 DW (dry weight)] and two exposure times of L. hexandra to CO= (d 180 and 360). A total of eight treatments with four replicates maintained at random locations in a microtunnel with an average temperature of 29 ± 6 °C and humidity at field capacity of 32 ± 5 %. The experimental unit was a plastic container with 4 kg of dry soil and one L. hexandra plant.
Rhizobacterial population
Azospirillum bacteria were grown on Congo red agar(29), Azotobacter bacteria on Asby agar(29), and Pseudomonas bacteria on cetrimide + glycerol agar(30). Cultures were incubated at 28 °C for 72 h, and counts were expressed as colony forming units (CFU) per gram of soil.
Chemical composition of L. hexandra
Destructive sampling of plant tissue (leaves and stems) from young plants emerging from the main plant of L. hexandra that were 180 and 360 days old was performed, dried in a forced air oven at 60 °C for 72 h and ground for crude protein, neutral detergent fiber, acid detergent fiber, and lignin analysis(31,32).
Production analysis
The evaluation of young plants in the tillering stage, aerial dry matter, and root dry matter was similar to the procedure used by Orocio-Carrillo et al(33).
Statistical analyses
The data collected for all variables were subjected to an analysis of variance and a multiple comparison test of means with Tukey's method (P<0.05), as well as to Pearson's bivariate correlation, using the SAS v.9.4 statistical software(34).
Results
Rhizobacterial population
Table 2 shows the changes in the mean values of the three groups of rhizobacteria due to the effect of crude oil on the soil and to the exposure time. The highest density of Azotobacter [438 and 132 x 103 CFU g-1 dry rhizosphere (d.r.)] was found in soil with 90 g kg-1 of crude oil, where it was 106.6 % and 40.4 % higher than the control at d 180 and 360 respectively. However, the densities of Azospirillum and Pseudomonas decreased. In general, the effect of crude oil contamination increases in the population of Azotobacter (81.7 %) and decreases in those of Azospirillum and Pseudomonas by 36 and 47.7 %, respectively. However, the effect of the time of evaluation induced a positive response in the density of Azospirillum and Pseudomonas, being 1.9 and 23.9 times higher at d 360 than at d 180.
Table 2: Changes in Azospirillum, Azotobacter, and Pseudomonas in the rhizosphere of L. hexandra exposed to crude oil at 180 and 360 days
The symbol % (+) represents an increase, and % (–), a decrease of Azospirillum, Azotobacter, and Pseudomonas at 180 and 360 d with respect to the values of the control treatment (0 g kg−1 crude oil).
abcde Different lowercase letters within a column indicate statistically different values (P≤0.05, n=4). * Statistically higher.
Chemical composition of L. hexandra
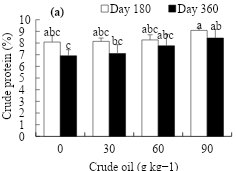
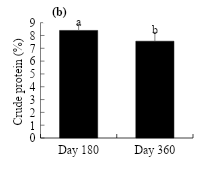
Figure 1 shows the tendency in crude protein content in the aerial dry matter of young L. hexandra plants by effect of crude oil dosage and exposure time. The highest crude protein content at d 180 (9.1 %) and 360 (8.4 %) was recorded at the 90 g kg-1 crude oil concentration, being 1 and 1.5 % higher than that of the control (Figure 1a). On the other hand, the evaluation time decreased from 8.4 to 7.5 % as the age of the plant increased from 180 to 360 d (Figure 1b).
Figure 1: Effect of the crude oil in the soil (a) and exposure time (b) on the crude protein percentage in L. hexandra
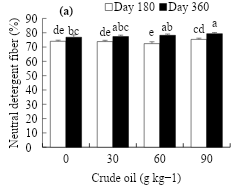
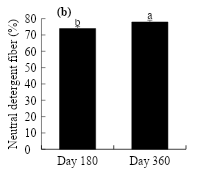
Figure 2 shows the performance in neutral detergent fiber. Significant differences were observed according to the oil dose and the evaluation time (P≤0.05). The dose of 90 g kg-1 of crude oil induced the highest neutral detergent fiber content at d 180 (75.3 %) and 360 (79.3 %), reaching an increase of 1.3 and 2.5 % over that of the control (Figure 2a). As for the evaluation time, it increased from 73.9 to 77.9 % as plant maturity increased from 180 to 360 d (Figure 2b).
Figure 2: Effect of crude oil in the soil (a) and the exposure time on the neutral detergent fiber in L. hexandra
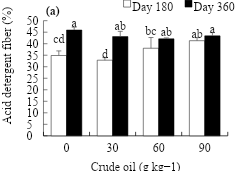
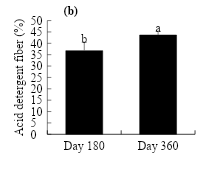
Figure 3 shows the behavior in the content of acid detergent fiber. At day 180, the dose of crude oil was observed to stimulate the acid detergent fiber, which was higher (41.3 %) in soil with 90 g kg-1 of crude oil, increasing 6.4 % over that of the control (Figure 3a). At day 360, crude oil concentrations showed a statistically similar response to the control. As for the evaluation time, the acid detergent fiber was stimulated to increase from 36.8 to 43.6 % at day 180 and 360, respectively (Figure 3b).
Figure 3: Effect of the crude oil on the soil (a) and time of exposure (b) on the acid detergent fiber in L. hexandra
abcd Different letters indicate different values (P≤0.05, n=4).
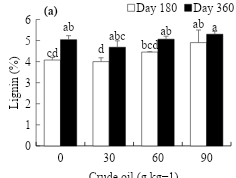
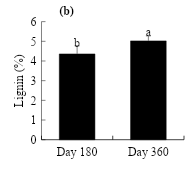
The effect of crude oil on the lignin content in L. hexandra harvested at different ages showed significant differences (P≤0.05) (Figure 4). At d 180, it is observed that high concentrations of oil stimulate lignin, being higher (4.9 %) in soil with 90 g kg-1 of crude oil, increasing 1 % with respect to the control (Figure 4a). On the other hand, the effect of evaluation time induced a positive response, reaching an increase of 4.3 to 5 % as plant age advanced (Figure 4b).
Figure 4: Effect of crude oil on the soil (a) and exposure time (b) on the percentage of lignin in L. hexandra
abcd Different letters indicate different values (P≤0.05, n=4).
L. hexandra production
The effects of crude oil doses and evaluation time on the means of young L. hexandra plants in the tillering stage and their aerial dry matter and root dry matter exhibited significant differences (P<0.05) (Table 3). Exposure to 90 g kg-1 of crude oil promoted an increase of up to 300 and 89.3 % of young plants in the tillering stage compared to the control soil at d 180 and 360 respectively. The results for root dry matter increased up to 203.7 and 169.7 % at d 180 and 360, respectively, in the most contaminated soil, with respect to the control soil. However, the aerial dry matter production of the grass decreased up to 25.9 and 18.9 % at day 180 and 360 with the highest dose. In general, the effect of crude oil contamination on young plants in the tillering stage increases by 78.6 % and radical dry matter 135.1 %. However, it decreased 13.4 % in the aerial dry matter, although this reduction was statistically equal to the control soil. The effect of the evaluation time induced a positive response in young L. hexandra plants in the tillering stage and the aerial and root dry matter, which were 210.8, 378.7, and 545 % higher at d 360 than at d 180.
Table 3: Changes in young plants in the tillering stage, the aerial dry matter and the root dry matter of L. hexandra exposed to crude oil at 180 and 360 days
The symbol % (+) represents an increase, and % (-) a decrease, of the aerial dry matter and root dry matter of young plants at d 180 and 360, compared to the values of the control treatment (0 g kg-1 of crude oil).
abcde Different lowercase letters within each column represent different values (P≤0.05, n=4). *Statistically higher.
Correlation of variables are presented in Table 4.
Discussion
Rhizobacterial population
Reports indicate that the rhizosphere of grasses used for the removal of total petroleum hydrocarbons from soils contaminated with crude oil host intense microbial activity, including that of plant growth-promoting rhizobacteria(35,36). The significant increase in the Azotobacter population (0.361*) (Table 2 and 4) in the rhizosphere of L. hexandra is similar to that reported in other studies evaluating the same grass exposed to 60-180 g kg-1 of total petroleum hydrocarbons(20) and Echinochloa polystachya K. exposed to 65.89 g kg−1 of total petroleum hydrocarbons(37). The increase in the Azotobacter population could be due to the adaptation of the bacterium through the secretion of extracellular enzymes essential for the initial degradation of high molecular weight substrates(38) such as petroleum hydrocarbons. Similarly, it may be an adaptation to a reduction in the availability of essential nutrients such as nitrogen due to the properties of crude oil, which tends to agglomerate in the soil(4;5). In addition, it has been shown that under stress conditions plants increase root exudation(39), which become a source of nutrients and stimulating substances for the growth of microorganisms(40). However, not all microorganisms can adapt quickly, probably because of the toxic hydrocarbons in crude oil, which make it difficult for some microbial species to grow and survive(41). In addition, the presence of oil can modify the physicochemical properties of the soil(4), generating unfavorable conditions for microorganisms(42). In contrast, the current study revealed that the population of Azospirillum and Pseudomonas decreased with increased crude oil, demonstrating the negative effects of this pollutant; at the same time, with the extension of time, the stimulation of both populations was promoted (0.778**, 0.767**), indicating that the bacteria need some time to acclimatize and achieve significant growth. Similar results were reported in other investigations(1,7), where they found a significant increase of rhizobacteria over time in the rhizosphere of L. hexandra and Urochloa brizantha Hochst exposed to oil.
Chemical composition of L. hexandra
The present study showed increases in crude protein content in shoots of L. hexandra planted in soil with crude oil (0.551**) relative to the control. At d 180 and 360 at doses of 60 and 90 g kg-1 of crude oil, the tendency is for the crude protein to increase with respect to the control, while at doses of 30 g kg-1 there was no statistical difference (Figure 1a). A similar effect was reported by Orocio-Carrillo et al(13), who found an increase in crude protein in L. hexandra exposed to 102 g kg-1 of crude oil. Likewise, the protein content in Simmodsia chinensis L. and Vigna unguiculata L. respectively has reportedly(43,44) increased due to the effect of crude oil in soil. Roa et al(45) report an increase in protein concentration in Triticum aestivum L. due to the effect of sulfur fertilization. Therefore, the increase in protein in this study could be due to the nitrogen and sulfur content present in the crude oil(46). On the other hand, crude protein is diluted as the age of the grass increases (−0.497**). It has been mentioned(47) that the increase in the age of the grass results in a decrease in protein, as dry matter production increases. In this regard, this study found a negative and highly significant relationship between crude protein and aerial dry matter (−0.564**). As the maturity stage of grasses increases, so does the content of structural carbohydrates and lignin, while the protein content decreases(48).
This study shows that the neutral detergent fiber at d 180 did not change in contaminated soils with respect to the control; however, at d 360, the tendency was to increase very slightly with high doses of crude oil (60 and 90 g kg-1), but at 30 g kg-1 there was no statistical difference compared to the control. On the other hand, neutral detergent fiber increased due to the effect of the evaluation time (0.832**). Similar tendencies were reported by other researchers(49,50), who evaluated the percentages of neutral detergent fiber of fountaingrass at different harvesting ages and observed an increase in its concentration of 5.5 and 17.9 % between day 30 and 167 d, respectively. It has also been mentioned(51) that, as forage maturity increases, the concentrations of neutral detergent fiber in stems and leaves augment, reducing the voluntary consumption of forages.
The concentration of acid detergent fiber in this study shows that it increased at high doses of oil (60 and 90 g kg-1) with respect to the control at d 180, but at doses of 30 g kg-1 the response was not statistically different from that of the control. At d 360, the tendency was to decrease at doses of 60 and 90 g kg-1, while at doses of 30 g kg-1 there were no statistical differences in relation to the control. On the other hand, the concentration of acid detergent fiber augmented as the age of the grass increased (0.741**). Similar values were reported by Schnellmann et al(52) when evaluating the nutritional quality of Megathyrsus maximus Jacq., as they recorded values of 29.3 % at d 90 and 34.4 % at d 180. Similarly, Álvarez-Vázquez et al(50) found an increase of 40.36 % at d 33 and 58.5 % at d 180 when evaluating the chemical composition of Cenchrus sp. grass. Acid detergent fiber is reported to be an important component that regulates forage quality and is positively related to the crop’s age or stage of development, with forage quality declining as fiber becomes a predominant component(53).
In the present study, one explanation for the increase in high doses of crude oil may be the mechanism of osmotic adjustment that favors the accumulation of compatible solutes, which are organic compounds that do not interfere with cell metabolism, even at high concentrations, and can act as antioxidants to minimize the impact of abiotic stress on the plant(24). Several studies have shown that lignin increases in response to various environmental stresses(26,27), playing a role in the adaptation of plants to their environment(54). On the other hand, the lignin content is stimulated (0.661**) as the age of maturity of the plant increases. Rosales and Pinzón(48) mentioned that, as the grass maturity stage increases, the proportion of cell wall components, including lignin, increases, reducing the digestibility of the grass.
L. hexandra production
In pastures, the addition of crude oil to the soil has been reported to induce a significant reduction of plant dry matter(55). It should be noted that, at d 180, aerial dry matter production was reduced only by 2.9 % at a concentration of 30 g kg-1 of crude oil, compared to the control, demonstrating the high tolerance of the grass in these conditions; however, as the dose of crude oil in the soil increases (60 and 90 g kg-1), the percentage reduction increases both at d 180 and at d 360 with respect to the control. The negative impact of crude oil on the aerial dry matter of L. hexandra has also been reported(13,56). Crude oil leads to negative changes in soil properties such as reduced moisture absorption and retention capacity, water repellency and insufficient soil aeration(57,58), which consequently makes plant growth even more difficult. On the other hand, the production of young plants in the tillering stage and root dry matter were stimulated (0.465**, 0.362*). This behavior was also mentioned in other researches(13,56). The increase in these variables may indicate a strategy of L. hexandra to survive and adapt to soil contamination by crude oil. The increase of young plants in the tillering stage and root dry matter in oiled soils could be explained by the increase in soil organic carbon content due to the degradation of crude oil(18). Likewise, the increase in roots could be due to a response of reinforcing the root tissues in order to limit soil nutrient deficiency stress induced by crude oil(5). Studies have reported increased root biomass in soils treated with petroleum hydrocarbons(22).
Conclusions and implications
The effect of high doses of crude oil in the soil induces in the rhizosphere of L. hexandra high populations of bacteria of the Azotobacter group; however, it caused the inhibition of Azospirillum and Pseudomonas. On the other hand, crude oil is positively correlated with crude protein and lignin synthesis, playing a role in grass adaptation to its environment. Likewise, high doses of crude oil induced a positive response in the production of young plants in the tillering stage and root dry matter, which indicates a strategy developed by the grass to survive and adapt to oil contamination. However, the chemical composition of the grass was affected by the age of harvest, in which the percentage of neutral detergent fiber, acid detergent fiber, and lignin increased, while the concentration of crude protein decreased. Therefore, even if the grass is able to adapt to the stress induced by crude oil, its protein content and, therefore, its nutritional value are affected, which can be detrimental to farmers and cause long-term deterioration of the affected property.
Acknowledgments
José Alberto Orocio Carrillo is grateful to the National Council for Humanities, Sciences, and Technologies (Consejo Nacional de Humanidades, Ciencias y Tecnologías, CONAHCYT) of Mexico for the scholarship No. 813712, granted his doctoral studies in Agricultural Sciences in the Tropic.
Conflict of interest
The authors declare that they have no conflict of interest.
Literature cited:
Table 4: Correlation of variables
CO= crude oil; PJM= young plants in the tillering stage; ADM= aerial dry matter; RDM= root dry matter; CP= crude protein; NDF= neutral detergent fiber; ADF= acid detergent fiber; AZP= Azospirillum; AZT= Azotobacter; PSE= Pseudomonas.
* P≤0.05. ** P≤0.01. NS= non significant.