https://doi.org/10.22319/rmcp.v16i1.6733
Artículo
Nivel dietético de concentrado de proteína de papa y su efecto sobre la concentración intestinal de citocinas y ácidos grasos volátiles en lechones destetados
Konisgmar Escobar-García a
Tércia Cesária Reis de Souza a
Mauricio Díaz-Muñoz b
Samantha Elizabeth Bautista-Marín a*
a Universidad Autónoma de Querétaro. Facultad de Ciencias Naturales, Licenciatura en Medicina Veterinaria y Zootecnia, Av. de las Ciencias s/n, Juriquilla. 76230, Querétaro, México.
b Universidad Nacional Autónoma de México. Instituto de Neurobiología. Querétaro, México.
* Autor de correspondencia: samantha.bautista@uaq.mx
Resumen:
El destete es un evento traumático para el lechón, ya que implica cambios que son responsables de alteraciones gastrointestinales, una disminución de la productividad o incluso la muerte. El desarrollo de resistencia bacteriana debido a la inclusión de dosis subterapéuticas de antibióticos en las dietas de inicio se ha convertido en un importante asunto de salud pública, prohibiendo su inclusión en la alimentación animal. El concentrado de proteína de papa (CPP) se ha considerado una alternativa para regular la inflamación y los trastornos intestinales debido a su contenido de péptidos antimicrobianos que tienen efectos beneficiosos sobre la homeostasis intestinal. En este estudio se evaluó el efecto del nivel de inclusión de CPP en una dieta libre de antibióticos sobre la concentración de marcadores inflamatorios como interleucina-12p40 (IL-12p40) y factor de necrosis tumoral alfa (TNF-α) en el tejido ileal y la concentración de ácidos grasos volátiles (AGV) en la digesta colónica. Se asignaron 90 lechones a tres tratamientos: 1, dieta basal (C) (dieta sin antibióticos ni CPP); 2, dieta basal con 6 % de CPP (CPP 6%) y 3, dieta basal con 8 % de CPP (CPP 8%). A los 15 días posdestete, se eutanasiaron seis lechones por tratamiento para la recolección de muestras. El grupo CPP 8% tuvo los niveles más altos de AGV y la concentración más baja de citocinas inflamatorias en comparación con el grupo C, que tuvo los niveles más bajos de AGV y la concentración más alta de marcadores inflamatorios. La inclusión de CPP en las dietas de inicio de lechones destetados puede ser una alternativa eficaz para regular la disbiosis intestinal durante el destete.
Palabras clave: Proteínas de papa, Citocinas proinflamatorias, Fermentación intestinal, Destete.
Recibido: 28/06/2024
Aceptado: 23/10/2024
Introducción
El desarrollo celular gastrointestinal, la fisiología, la microbiota y la inmunidad ajustan sus funciones de acuerdo con los requerimientos del animal, para nutrir y conferir protección contra microorganismos potencialmente patógenos que podrían comprometer la supervivencia de los lechones destetados precozmente, esto debido a la inmadurez de su actividad digestiva e inmunológica(1,2). El destete de los lechones se asocia a un desequilibrio entre las respuestas inmunitarias e inflamatorias, lo que lleva al desarrollo de enfermedades por la secreción de citocinas proinflamatorias(3). Las citocinas son un grupo de proteínas y glicoproteínas sintetizadas por diferentes líneas celulares, responsables de la regulación de las respuestas inflamatorias e inmunitarias que actúan a nivel sistémico modulando la actividad celular a través de la interacción con receptores de membrana específicos que desencadenan una cascada de señalización. La síntesis y liberación de citocinas puede ser estimulada por una incorrecta activación inflamatoria causada por el alimento, el medio ambiente, las bacterias y los metabolitos, así como por la presencia de otras citocinas proinflamatorias, incluyendo la interleucina-1 (IL-1), el factor de necrosis tumoral alfa (TNF-α) y la interleucina-6 (IL-6), que inducen la producción de proteínas de fase aguda(4,5). La citocina proinflamatoria TNF-α actúa como un mediador de la inflamación aguda que aumenta la producción de IL-1 e IL-6(6).
La microbiota intestinal es una población compleja con cientos de microorganismos diversos que contribuyen a la descomposición de los nutrientes y al metabolismo energético, afectando al sistema inmunitario y a la homeostasis(7). La microbiota del intestino grueso es la responsable de la fermentación de los carbohidratos y proteínas no digeribles en las dietas de los lechones para producir una serie de metabolitos denominados ácidos grasos volátiles (AGV). Entre los diversos metabolitos, los ácidos grasos de cadena corta (AGCC) han recibido una amplia atención debido a sus efectos positivos sobre la salud(8).
Los antibióticos han sido ampliamente utilizados en la industria porcina como promotores del crecimiento, lo que ha llevado al desarrollo de resistencia bacteriana y a la presencia de residuos de antibióticos en productos animales(9). Para enfrentar esta problemática se ha planteado la necesidad de identificar alternativas que reemplacen el uso de antibióticos manteniendo los parámetros productivos de crecimiento. El concentrado de proteína de papa ha sido considerado como una valiosa fuente de aminoácidos esenciales que podría reemplazar la inclusión de proteína animal en las dietas iniciadoras de lechones debido a la presencia de péptidos antimicrobianos (PAM) y el contenido de proteína de alta digestibilidad(10,11). La actividad antibacteriana de los PAM del CPP se basa principalmente en la interacción de péptidos cargados positivamente con componentes cargados negativamente de la membrana bacteriana, como fosfolípidos y ácidos teicoicos de bacterias grampositivas o lipopolisacáridos de bacterias gramnegativas, lo que conduce a la formación de poros, permeabilización de la membrana y lisis celular después de la relocalización en la membrana citosólica(12).
La actividad antimicrobiana del CPP podría ser una alternativa interesante al uso de antibióticos como promotores del crecimiento, alterando la composición de la microbiota para reducir la competencia por nutrientes, reducir el patógeno y controlar la mucina; sin embargo, aún no se ha determinado el nivel de inclusión para la nutrición de los lechones, por lo tanto, es necesario evaluar el efecto de diferentes niveles de inclusión.
El objetivo de este estudio fue establecer el nivel óptimo de inclusión de CPP para disminuir el proceso inflamatorio posdestete, definido por la presencia de marcadores proinflamatorios, así como el perfil de AGV.
Material y métodos
Manejo de animales
Todos los procedimientos experimentales con animales se llevaron a cabo de acuerdo con los lineamientos de la Norma Oficial Mexicana NOM-062-ZOO-1999 para la producción, cuidado y uso de animales para experimentación(13) y los lineamientos de los Principios Rectores Internacionales para la Investigación Biomédica con Animales(14). El diseño experimental y los procedimientos de este estudio fueron revisados y aprobados por el Comité de Bioética de la Facultad de Ciencias Naturales de la Universidad Autónoma de Querétaro (número de aprobación: 96FCN2021).
En este estudio se utilizaron noventa (90) lechones híbridos (Large white x Landrace x PIC337) destetados a los 21 ± 2 días y con un peso de 6.85 ± 0.93 kg, y se asignaron a tres dietas experimentales según el origen de la camada y el peso corporal, y se alojaron en seis corrales por dieta, cada uno con cinco lechones, haciendo un total de 30 lechones por dieta. Se formularon tres dietas experimentales con una dieta basal sin antibióticos ni concentrado de proteína de papa (C), posteriormente se adicionó 6 % y 8 % de concentrado de proteína de papa a la dieta basal para establecer las otras dos dietas, a saber, CPP 6% y CPP 8%, respectivamente, como se describe en el Cuadro 1. Los corrales se equiparon con un comedero con seis espacios y bebederos de chupón en una sala de destete ambientalmente controlada (32 °C con -2 °C durante la primera y segunda semana posdestete).
Cuadro 1: Ingredientes y composición química porcentual de las dietas experimentales
*L-glutamina, ácido L-glutámico (1:1).**vitaminas por kg de dieta, A: 13,000 UI, E: 160 mg, K: 9 mg, tiamina: 4 mg, riboflavina: 12 mg, piridoxina: 6 mg, cianocobalamina: 0.07 mg, niacina: 66 mg, pantotenato: 46 mg, folato: 5 mg, biotina: 0.67 mg, C: 266 mg. ***minerales por kg de dieta, manganeso: 32 mg, zinc: 120 mg, hierro: 100 mg, cobre: 12 mg, yodo 0.8 mg, selenio: 0.25 mg, cobalto: 0.6 mg.
a Valor analizado, b Valor calculado.
Colecta de muestras
A los 15 días después del destete, se practicó la eutanasia a seis lechones por grupo de dieta para su posterior muestreo. Los animales se tranquilizaron con 20 mg/kg de azaperona (Sural® Chinoin, Ciudad de México, México) y luego sacificados con una sobredosis de pentobarbital sódico (Pisabental®, PiSA Agropecuaria, Hidalgo, México). Se tomaron muestras de secciones de tejido ileal (10 cm) desde 5 cm antes de la válvula ileocecal, se lavaron suavemente con agua destilada y luego se diseccionaron y fijaron en una solución de formalina tamponada al 10 %. Las secciones ileales se incrustaron en parafina y se realizaron cortes de 5 μm de grosor. El contenido de la digestión colónica se colectó, congeló y almacenó para su análisis posterior.
Análisis de citocinas
Los cortes de 5 μm se utilizaron para cuantificar TNF-α e IL-12p40 mediante una técnica de inmunofluorescencia adaptada de la técnica empleada por Bautista-Marín et al(15). Los cortes de íleon se desparafinaron durante 24 h a 60 °C en una estufa de calor seco y luego se rehidrataron en xilol al 100 % (10 min), etanol al 100 % (5 min), etanol al 96 % (5 min) y agua desionizada (10 min), en este orden. A continuación, los cortes de íleon se remojaron en concentrado tampón de citrato sódico salino (Sigma Aldrich, EE. UU.) en un baño de agua a 80 °C durante 25 min. A continuación, los cortes de íleon se bloquearon en leche descremada al 1% durante 1 h, se lavaron tres veces durante 5 min cada uno con solución salina tamponada con TRIS al 0.05% más Tween (TBST) y se incubaron durante 2 h a 4 °C con los siguientes anticuerpos diluidos en TBST: i) anti TNF-α de conejo (ab 6671) (Abcam, Cambridge, MA, EE. UU.) a una dilución 1:200, ii) anti IL-12p40 de conejo (ab 106270) (Abcam, Cambridge, MA, EE. UU.) a una dilución 1:100 e incubado durante 16 h. A continuación, los cortes de íleon se lavaron tres veces durante 5 min cada uno con TBST al 0.05 % y se incubaron durante 2 h con el anticuerpo secundario Alexa Fluor 488 pollo anticonejo IgG (H+L) (A-21441) (Invitrogen Molecular Probes Inc., Eugene, OR, EE. UU.) a una dilución de 1:500. La intensidad de la fluorescencia [unidades arbitrarias de fluorescencia (UAF)] se evaluó con los controles positivos (anticuerpos primarios y secundarios) y negativos (solo anticuerpos secundarios) considerando el mismo tamaño de área en el tejido ileal en ambas condiciones. Para el análisis, la fluorescencia se visualizó y midió con un microscopio de fluorescencia (Eclipse E600, Nikon) e Image-Pro Plus versión 6.1 en 10 vellosidades intestinales por lechón y se fotografiaron con un aumento de 40x y luego se seleccionaron 10 secciones de cada vellosidad intestinal por foto para obtener 100 mediciones por lechón.
Análisis de ácidos grasos volátiles
Se pesaron aproximadamente 2 g de contenido de digesta colónica y se colocaron en tubos de centrífuga previamente identificados, después se agregaron 3 ml de agua de grado HPLC a los tubos y se mezclaron con un vórtex durante 30 seg. A continuación, los tubos se colocaron en una centrífuga Beckman Coulter a 4 °C durante 30 min. Con la ayuda de una jeringa de 3 ml, se extrajo el sobrenadante y se vació en un tubo de color ámbar después de pasar a través de un filtro de jeringa de 0.2 μm. Las concentraciones de ácidos acético, propiónico, butírico y valérico (AGCC); los ácidos isobutírico, isovalérico e isocaproico (AGR) se analizaron en un cromatógrafo de gases Agilent 6890 equipado con un detector de ionización de llama y una columna DB-FFAP (30 m × 0.25 mm × 0.25 μm; Agilent Technologies, Wilmington, DE). Se utilizaron los siguientes parámetros del cromatógrafo de gases: modo dividido, 20:1; temperatura de entrada, 220 °C; presión de entrada inicial, 168 kPa; volumen de inyección, 1 μl; caudal constante de la columna (He), 1.4 ml/min; y la temperatura del detector, 250 °C.
Análisis estadístico
La concentración de marcadores inflamatorios, así como la concentración de ácidos grasos volátiles se analizaron utilizando un diseño completamente al azar, cada lechón se consideró como una unidad experimental(16). Se utilizó el ANOVA para analizar las diferencias entre los grupos de dietas. Las medias se compararon con la prueba de Tukey utilizando el procedimiento GLM de SAS y las diferencias se consideraron estadísticamente significativas con P<0.05(17).
Resultados
Concentración de citocinas inflamatorias
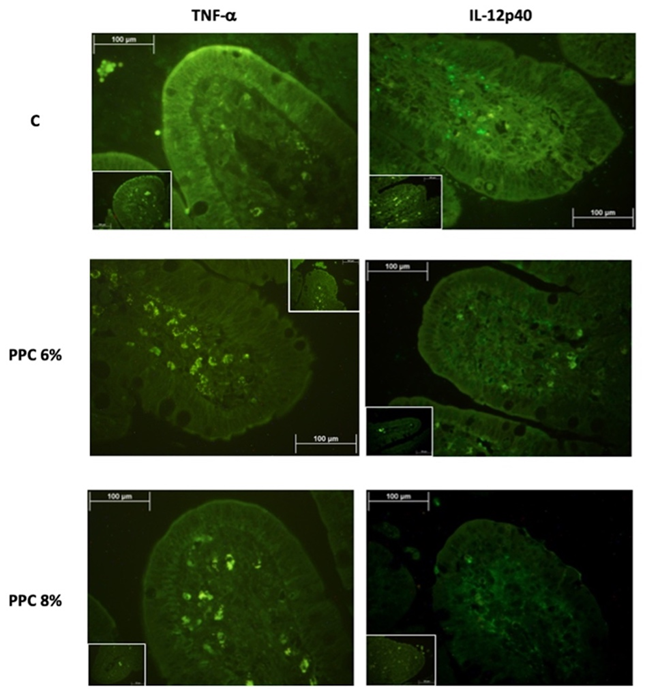
Las concentraciones de TNF-α e IL-12p40 en las vellosidades ileales de lechones destetados se vieron afectadas (P<0.0001) por la dieta (Cuadro 2). Los lechones alimentados con la dieta C tuvieron las concentraciones más altas de todos los marcadores inflamatorios. Los animales alimentados con la dieta CPP 6% mostraron concentraciones intermedias, y el grupo CPP 8% tuvo las concentraciones más bajas (P<0.0001). Se tuvo la señal más alta de UAF para IL-12p40 en comparación con TNF-α en los lechones alimentados con todas las dietas experimentales, pero particularmente en aquellos alimentados con la dieta C (Figura 1).
Cuadro 2: Efecto del nivel de inclusión de concentrado de proteína de papa (CPP) sobre la concentración de citocinas proinflamatorias en íleon
UAF= unidades arbitrarias de fluorescencia; CPP= concentrado de proteína de papa; C= dieta sin antibióticos ni CPP; CPP 6%= dieta basal con 6 % de CPP; CPP 8%= dieta basal con 8 % de CPP; P= probabilidad. EEM= error estándar de la media
abc Las medias de las filas con diferentes superíndices difieren significativamente en P<0.05. ***P<0.0001.
Figura 1: Concentración de TNF-α e IL-12p40 en el íleon de lechones alimentados con tres dietas experimentales empleando la tinción de inmunofluorescencia
Los cuadros representan los controles sin el anticuerpo primario.
Concentración de ácidos grasos volátiles
Los perfiles de fermentación en el colon se vieron afectados por la dieta (Cuadro 3). Los lechones alimentados con la dieta C presentaron una menor concentración de AGV totales (P<0.0001) que los lechones alimentados con dietas con CPP al 6 y 8 %. En consecuencia, la inclusión de CPP en ambos niveles en la dieta aumentó la producción de AGCC y ácidos grasos ramificados (AGR) en el colon en comparación con la dieta C (P<0.0001). Los animales alimentados con la dieta CPP 8 % tuvieron una mayor concentración de AGCC (P<0.0001) que los animales alimentados con la dieta CPP 6 % o la dieta C. El ácido acético fue el AGCC más abundante en la digesta de todas las dietas experimentales, seguido por el butírico y el propiónico. Se observaron mayores concentraciones de AGCR en la digesta de los animales alimentados con la dieta CPP 8% que en los alimentados con las dietas CPP 6% y C (P<0.0001).
Cuadro 3: Efecto del nivel de inclusión de concentrado de proteína de papa (CPP) sobre la producción de ácidos grasos volátiles en el colon
C= dieta basal sin antibiótico ni CPP; CPP 6%= dieta basal con 6 % de CPP; CPP 8%= dieta basal con 8 % de CPP; AGR= ácidos grasos ramificados; AGCC= ácidos grasos de cadena corta.
P= probabilidad estadística; EEM= error estándar de la media.
abc Las medias de las filas con diferentes superíndices difieren significativamente en P<0.05. ***P<0.0001.
Discusión
Los resultados observados en trabajos anteriores sugieren que los marcadores inflamatorios en el íleon, como son las citocinas, juegan un papel central en la inmunidad celular y participan en el mantenimiento de la integridad del tejido. Se esperaban cambios en las concentraciones de citocinas intestinales para los lechones destetados, debido a cambios en los componentes dietéticos y a los factores ambientales estresantes durante este período, que conducen a importantes adaptaciones morfofisiológicas en el tracto gastrointestinal(1,4,5). Los antígenos microbianos presentes en la superficie intestinal generan una respuesta inflamatoria temprana mediante la secreción de citocinas proinflamatorias, que activan una vía de señalización, que conduce a la liberación y translocación al núcleo del factor nuclear-κB (NF-κB), que sirve como factor de transcripción para la síntesis de IL-6 y TNF-α(18-20).
La concentración de interleucina 12 subunidad p40 (IL-12p40) surge cuando ocurre un proceso inflamatorio crónico en el tracto gastrointestinal(21). La inflamación modulada por la IL-12p40 es desencadenada a través la Janus quinasa (JAK/STAT), la cual es fosforilada, translocándose al núcleo donde se transcribe la IL-12p40(22).
La conjunción entre las células epiteliales intestinales y las células de la lámina propia subyacente transfiere señales relacionadas con el sistema inmunitario a la inmunidad adaptativa local, que posteriormente ayudan a mantener la homeostasis inmunitaria intestinal(23).
La alteración de la integridad de la mucosa intestinal genera un proceso inflamatorio que activa el NF-κB y la consecuente producción de citocinas proinflamatorias(6). La respuesta proinflamatoria se exacerba cuando los lechones destetados precozmente son alimentados con dietas libres de antibióticos(24).
La suplementación dietética con concentrado de proteína de papa ayudó a regular la síntesis de los marcadores inflamatorios TNF-α e IL-12p40, cuya concentración disminuyó en 43.31 % y 60.45 %, respectivamente, en la dieta CPP 8% en comparación con la dieta C. La regulación negativa de las citocinas por los péptidos antimicrobianos del CPP se confirmó con los resultados actuales, ya que la concentración de TNF-α e IL-12p40 disminuyó en los lechones alimentados con la dieta CPP 6% y CPP 8%. El efecto antimicrobiano sobre la población bacteriana del colon probablemente disminuyó la vía de señalización que induce la liberación de NF-κB(19), que fue altamente marcada por el anticuerpo primario utilizado en el presente estudio de inmunofluorescencia (Figura 1). El LPS es un componente principal de las membranas externas de las bacterias gramnegativas y puede ser reconocido por el receptor tipo Toll 4 (TLR4) activando la producción de citocinas proinflamatorias por parte de las células inmunitarias(25). Las funciones antiinflamatorias del CPP se deben principalmente a la actividad neutralizante de LPS, que suprime las vías de señalización de TLR4, como la proteína quinasa activada por mitógenos [MAPK] y la señalización de NF-κB(26). Además, la inclusión de CPP en la dieta disminuyó la síntesis de IL-12p40 y, lo que se atribuye a la falta de estímulo generado por microorganismos patógenos y a la inhibición de la vía mediada por JAK/STAT(21,22). Por lo tanto, la suplementación dietética con CPP regula la respuesta inflamatoria e indirectamente protege la mucosa intestinal, disminuyendo los efectos perjudiciales del destete sobre las células intestinales(27,28).
Los ácidos grasos volátiles también tienen un papel clave en la regulación de los mecanismos inflamatorios que confieren un efecto protector al modular la liberación de citocinas inflamatorias y el reclutamiento de células inmunes a través de receptores de superficie o por la inhibición de la actividad enzimática(29). Los AGCC se definen como grupos de ácidos grasos volátiles que comprenden menos de seis carbonos, principalmente acetato, propionato y butirato, que se sabe que son la principal fuente de energía para los colonocitos y son esenciales para mantener el metabolismo normal de la mucosa del colon, incluyendo el crecimiento y la proliferación de colonocitos(2). Estos ácidos hacen posible la regulación del pH del contenido colónico, inhibiendo el crecimiento y desarrollo de bacterias potencialmente patógenas como E. coli, Clostridium y Salmonella. Los AGV estimulan la secreción luminal de bicarbonato, que tiene un importante efecto tampón sobre la regulación del pH luminal, al tiempo que induce la absorción de sodio y agua a nivel de la mucosa(30).
El aumento de AGCC observado en lechones alimentados con CPP 6% y CPP 8%, en comparación con los producidos por lechones alimentados con una dieta sin antibiótico ni CPP, puede ser probablemente el resultado de alteraciones ambientales intestinales que favorecen el establecimiento de bacterias beneficiosas y limitan el crecimiento de microbios patógenos(31). A su vez, el ácido acético puede ser metabolizado a ácido butírico, el cual aporta entre el 70 % y el 90 % de la energía necesaria para el metabolismo de los colonocitos(31). Estos cambios pueden reducir la incidencia y la gravedad de la diarrea posdestete y mejorar el crecimiento de los lechones(26). Los AGCC promueven un ambiente intestinal eubiótico al mantener un pH ligeramente ácido, lo que suprime el crecimiento de bacterias potencialmente patógenas que requieren un pH alcalino para sobrevivir(32). La microbiota beneficiosa prolifera a través de la exclusión competitiva, ocupando sitios de unión en la mucosa intestinal que, de otro modo, podrían ser empleados por bacterias patógenas(33). El establecimiento de microorganismos benéficos modula el desarrollo de bacterias patógenas a través de la secreción de bacteriocinas, que destruyen la pared celular bacteriana imitando el mecanismo de acción de algunos antibióticos(32,33). Los AGCC podrían actuar como inhibidores de la histona deacetilasa (HDAC) y, por lo tanto, atenuar las respuestas inflamatorias al bloquear la señalización del factor nuclear-κB (NF-κB)(34). Además, los AGCC también tienen un efecto directo sobre los ligandos del receptor de hidrocarburos de arilo (AHR) derivados de la microbiota, que potencian las actividades inmunosupresoras de las células T reguladoras (Treg) durante la inflamación, e influyen indirectamente en el reclutamiento y la activación de los neutrófilos(23). El aumento de la producción de ácidos acético, propiónico y butírico está probablemente relacionado con la modulación de la población microbiana intestinal a través de la actividad antimicrobiana del CPP.
La influencia de los factores dietéticos, a través de intervenciones dietéticas como el ácido butírico y las bacterias productoras de ácido, puede aumentar los AGCC en el intestino; sin embargo, un exceso de AGCC en el intestino grueso podría promover el desarrollo del síndrome metabólico a través del eje microbiota intestinal-cerebro-célula β(34). Una cantidad adecuada de AGCC en el intestino puede ser beneficiosa para la salud intestinal de los lechones destetados precozmente. El contenido y la proporción de AGV en el intestino se ven afectados por las características de las proteínas, como la solubilidad y la fermentabilidad(35). En cuanto a la fermentación de proteínas, la proteólisis es el primer paso en la utilización de proteínas por parte de las bacterias, seguido por la desaminación y la descarboxilación de los aminoácidos, lo que limita su disponibilidad para el hospedero y produce varios compuestos putrefactos como amoníaco, aminas, ácidos grasos ramificados, indoles, fenoles y compuestos que contienen azufre(36). Estos compuestos pueden tener efectos tóxicos en los lechones y pueden afectar a la función y la diversidad de la microbiota intestinal(37). Por lo tanto, puede haber un umbral para los AGCC y los AGR en el intestino, a pesar de que el CPP 8% tenía los niveles más altos de AGV totales, en particular los AGCC, también fue la dieta con el nivel más alto de AGR, lo que sugiere que el CPP 6% puede ser el nivel óptimo de inclusión, con el fin de prevenir efectos nocivos sobre la salud intestinal, manteniendo las propiedades inmunomoduladoras y antiinflamatorias de la inclusión de CPP.
Los resultados actuales comprueban la hipótesis de que el concentrado de proteína de papa puede modular los niveles de citocinas intestinales y aumentar la producción de AGCC. Los lechones alimentados con dietas suplementadas con CPP presentaron los niveles más bajos de marcadores inflamatorios que pueden ser comparables con los de los lechones alimentados con dietas con antibióticos(15). El efecto inmunomodulador del CPP a nivel intestinal se debe probablemente a la inhibición de vías de señalización como NF-κB y el complejo JAK/STAT que suprimen la síntesis de citocinas proinflamatorias(38). Un beneficio adicional observado con la inclusión de CPP en dietas libres de antibióticos fue el control de la inflamación crónica asociada al aumento de IL-12p40(39). La eficacia del CPP observada en el presente estudio tuvo un efecto positivo en el control de la inflamación intestinal aguda y crónica.
Conclusiones e implicaciones
La inclusión de concentrado de proteína de papa en dietas sin antibióticos puede mejorar la salud intestinal en el periodo posdestete. Además, el 6 % y el 8 % de CPP ayudaron a disminuir la respuesta inflamatoria de los marcadores proinflamatorios medidos, principalmente en la concentración de IL-12p40, así como probablemente ayudaron a establecer un entorno intestinal óptimo para la fermentación microbiana beneficiosa y la producción de AGCC, teniendo una influencia potencial en la proliferación de bacterias intestinales. No obstante, el CPP 6% puede ser el nivel óptimo de inclusión, previniendo los efectos nocivos de AGR sobre la salud intestinal, manteniendo las propiedades inmunomoduladoras y antiinflamatorias de la inclusión de CPP. Por lo tanto, la inclusión de concentrado de proteína de papa puede utilizarse como un aditivo alimentario eficaz en dietas sin antibióticos. Deben tenerse en cuenta consideraciones económicas para la selección del nivel óptimo de inclusión de CPP en la dieta de los lechones por parte del productor. Las actividades antibacterianas de amplio espectro del CPP se han demostrado indirectamente, lo que los convierte en alternativas prometedoras a los antibióticos. Las propiedades inmunomoduladoras del CPP significan que probablemente tengan un rendimiento similar en comparación con los antibióticos en la industria porcina. Se necesitan investigaciones futuras para dilucidar su efecto preciso sobre la concentración de citocinas antiinflamatorias y la diversidad de la población intestinal microbiana para comprender completamente su mecanismo de acción.
Agradecimientos
Esta investigación fue financiada por el Fondo para el Desarrollo del Conocimiento (FONDEC-UAQ 2021) número de registro FNV202104. Los autores agradecen a Miguel Ángel Chávez Vargas, Dr. Christian Molina Aguilar y Berenice Caballero Flores por su apoyo técnico.
Conflicto de intereses
Los autores declaran no tener ningún conflicto de intereses.
Literatura citada: