https://doi.org/10.22319/rmcp.v16i1.6724
Artículo
Preferencias forrajeras del borrego cimarrón (Ovis canadensis, Shaw) en Baja California, México
Ángel Méndez-Rosas a
Fernando Isaac Gastelum-Mendoza a
Guillermo Romero-Figueroa a*
Israel Guerrero-Cárdenas b
Hiram Rivera-Huerta c
Enrique de Jesús Ruiz-Mondragón a
Rafael Paredes-Montesinos a
Luz Adriana Tapia-Cabazos a
a Universidad Autónoma de Baja California. Facultad de Ciencias. Carretera Transpeninsular 3917, Col. Playitas, Ensenada, Baja California, México.
b Centro de Investigaciones Biológicas del Noroeste. Baja California Sur, México.
c Universidad Autónoma de Baja California. Facultad de Ciencias Marinas. Baja California, México.
*Autor de correspondencia: gastelummendozaisaac@gmail.com
Resumen:
El propósito de esta investigación fue analizar la composición y preferencias alimentarias del borrego cimarrón en las Sierras Juárez y Santa Isabel durante la época húmeda y seca de 2021-2022. Para evaluar la disponibilidad de forraje Se implementaron 17 líneas de Canfield de 100 m de longitud. La composición dietética se determinó mediante microhistología de muestras fecales, mientras que la diversidad del forraje y la dieta se calcularon utilizando el índice de Shannon. La selección forrajera se evaluó con el índice de Ivlev. Se observó que las formas biológicas más comunes en el hábitat del borrego cimarrón fueron las arbóreas y arbustivas. En las Sierras Juárez y Santa Isabel, se identificaron 31 y 43 especies de plantas en su dieta, respectivamente; las arbóreas y arbustivas fueron las más consumidas. No se encontraron diferencias en la dieta entre épocas y sitios. Larrea tridentata e Hisbiscus denudatus fueron las más frecuentes en la dieta, mientras que las preferidas incluyeron Eriogonum inflatum, H. denudatus, Horsfordia newberryi, Justicia californica y L. tridentata. Estos resultados ofrecen información para establecer estrategias de conservación y manejo comunitario del borrego cimarrón en Baja California.
Palabras clave: Borrego silvestre, Cordillera Molina, Matomí, Matorral desértico, Sierra Santa Isabel, Sierra Juárez.
Recibido: 06/06/2024
Aceptado: 11/10/2024
Introducción
El borrego cimarrón del desierto (Ovis canadensis) es una de las dos especies de borregos silvestres con distribución natural en Norteamérica(1). Actualmente se encuentra en vida libre en las regiones áridas montañosas del suroeste de los Estados Unidos de América y del noroeste de México(2). No obstante, hasta la segunda mitad del siglo XIX su área de distribución natural se extendía hasta la región noreste de México y abarcaba parte de los estados de Chihuahua, Coahuila y Nuevo León(3). Este decremento poblacional se debe a la degradación de su hábitat, la cacería furtiva y la transmisión de enfermedades por el ganado doméstico(1,2). Como resultado, la legislatura mexicana ha catalogado a esta especie como sujeta a Protección Especial (Pr) desde el año 2010(4).
El borrego cimarrón desempeña un importante papel ecológico al influir directamente en la dinámica de la vegetación(5,6) y en el ciclo de nutrientes del ecosistema(7). Por ello, los esfuerzos de conservación se han enfocado en preservar su hábitat natural para promover el desarrollo y establecimiento de sus poblaciones(8). Para lograr este objetivo es necesario comprender el uso que hace de diferentes tipos de forraje, lo que permite entender su capacidad de adaptabilidad a las variaciones en la disponibilidad y calidad de éste(9). Asimismo, el conocimiento sobre la dieta también permite la identificación de las áreas clave de forrajeo de la especie, la cual es información importante para su conservación(8,9,10).
Se han realizado diversos estudios en Norteamérica sobre los hábitos alimentarios del borrego cimarrón(8,11,12) en los que, se le describe como un consumidor oportunista con una preferencia por arbustos y hierbas en función de su disponibilidad(6,8,9). Sin embargo, en la península de Baja California sus hábitos forrajeros han sido poco estudiados y la mayoría de los estudios se basan en la observación directa o en el análisis de contenido estomacal(13), los cuales, no son representativos del consumo alimentario total, pero son valiosos porque indican un consumo significativo de pastos en una región altamente árida. Por lo anterior, el objetivo del presente estudio fue identificar y comparar la composición de la dieta del borrego cimarrón mediante análisis microhistológicos en dos sistemas montañosos de Baja California, México.
Material y métodos
Descripción del área de estudio
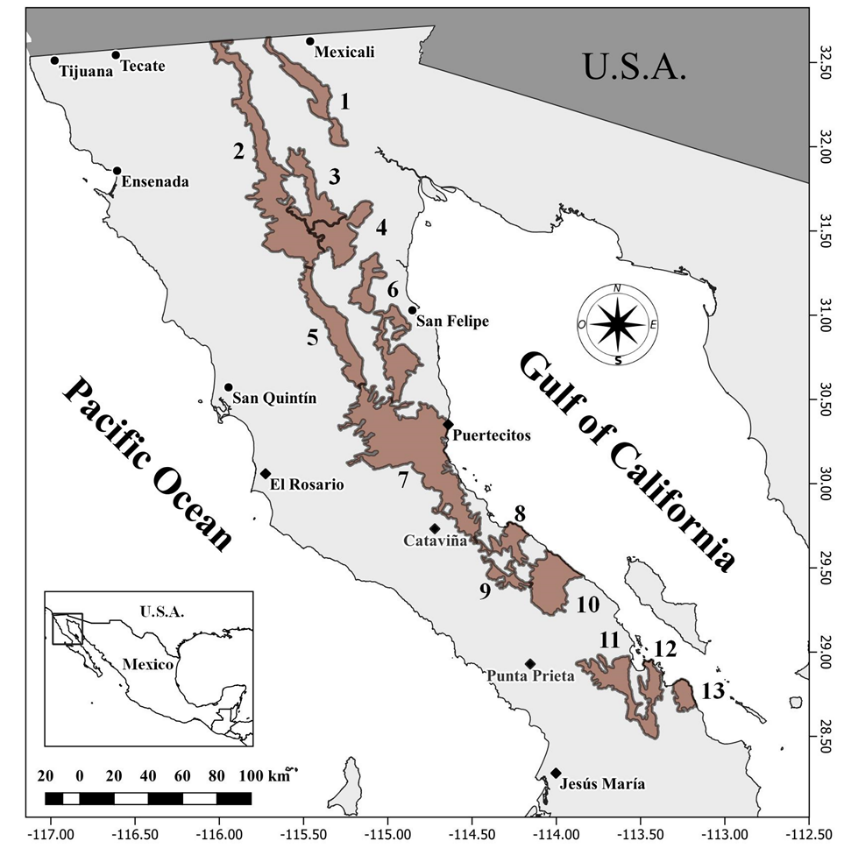
El estudio se realizó durante el periodo comprendido entre noviembre de 2021 y noviembre de 2022, en las cordilleras Sierra Juárez y Santa Isabel; en la región norte y centro del estado de Baja California, respectivamente (Figura 1). Estas áreas representan un hábitat continuo para el borrego cimarrón, ya que incluyen topografía accidentada, cañones y pendientes pronunciadas(14,15). Sierra Juárez tiene una extensión de 42,364 ha y se localiza entre las ciudades de Tecate y Mexicali; y Santa Isabel, abarca una superficie de 65,961 ha en la costa del Golfo de California, a 90 km al sur de la ciudad portuaria de San Felipe(15). Estas áreas pertenecen a la ecorregión Desierto de San Felipe, donde predomina el matorral desértico micrófilo, caracterizado por especies vegetales como la gobernadora (Larrea tridentata), la hierba del burro (Ambrosia dumosa), el ocotillo (Fouquieria splendens), el agave desértico (Agave deserti) y el palo fierro (Olneya tesota)(16). La temperatura media anual oscila entre los 12 y 22 °C. La precipitación promedio mensual oscila entre 0.0 mm y 0.8 mm, y se presenta durante todo el año, aunque los meses de invierno son los más húmedos(17).
Figura 1: Principales sistemas montañosos en el estado de Baja California, México (2. Sierra Juárez; 7. Sierra Santa Isabel)
Evaluación de la disponibilidad de forraje
La disponibilidad de forraje se evaluó mediante la frecuencia de cada especie vegetal por sitio y época del año (seca y húmeda). Para ello se utilizó el método de línea de intercepción o de Canfield. Para disminuir el error y aumentar la representación del muestreo, se generó una curva de acumulación para estimar el número de especies potenciales en cada ejido de acuerdo con los estimadores Jackknife(18) y Chao 2(19), mediante el paquete estadístico EstimateS V. 9.1.0. En total, se realizó la evaluación en 17 líneas de 100 metros de longitud de acuerdo a la curva de acumulación: en Sierra Juárez se establecieron cuatro líneas durante la estación seca (mayo y agosto de 2022) y cuatro durante la estación húmeda (noviembre de 2022); y en Sierra Santa Isabel se colocaron cuatro líneas en la época seca (abril y noviembre de 2022) y cinco en la época húmeda (junio y enero de 2022). En cada una se identificaron, contabilizaron y clasificaron todas las plantas que interceptaron la línea según su especie, cobertura lineal, altura y forma biológica (arbóreas y arbustivas, herbáceas, pastos y suculentas)(20). La distribución de las líneas se determinó con base en la información proporcionada por los miembros de cada ejido sobre las especies de plantas consumidas por el borrego cimarrón, la identificación de áreas con topografía asociada a la presencia del borrego cimarrón y en las observaciones directas generadas durante los recorridos de prospección realizados por el equipo del Laboratorio de Manejo y Conservación de Vida Silvestre de la Universidad Autónoma de Baja California. Los datos recopilados permitieron calcular la frecuencia de disponibilidad por especie vegetal, sitio de estudio y estación del año mediante la siguiente ecuación:
Disponibilidad de la especie i %=No. de líneas que contienen a la especie iNúmero total de líneas en el sitio x 100
Composición, diversidad y selección del forraje
Para identificar las especies que constituyen la dieta del borrego cimarrón se empleó la técnica microhistológica, la cual implica la identificación de patrones de estructuras celulares de la epidermis vegetal en muestras fecales(21). Se creó un catálogo de referencia que consistió en una colección de fotografías de estructuras celulares vegetales. Para ello, se recolectaron muestras de plantas que incluían flor, hojas y tallos en los mismos lugares donde se obtuvieron las muestras fecales. Las plantas recolectadas se prensaron y luego se identificaron taxonómicamente con la ayuda de la colección del herbario de la Universidad Autónoma de Baja California (BCMEX), la guía de plantas de Baja California(22), la plataforma Naturalista y consultas con expertos. Para elaborar el catálogo de referencia, las plantas se procesaron en un molino Wiley modelo Thomas tp4276 m004 con una malla de tamaño no. 20 (1 mm); posteriormente se aclararon con 20 ml de hipoclorito de sodio (cloro comercial) al 5 % y se montaron en portaobjetos. Las láminas (portaobjetos) se observaron en un microscopio digital de alta gama con un objetivo de 10X para identificar y fotografiar estructuras celulares diagnósticas: tricomas, estomas, cristales, arreglo de la epidermis, etc.(21)
Se realizó una recolección aleatoria de 195 muestras fecales frescas en ambas sierras durante las épocas seca y húmeda conforme a la metodología sugerida por Anthony y Smith(23). Previo al análisis histológico de las muestras fecales se tomaron cinco submuestras al azar en cada sitio y temporada de muestreo para formar muestras compuestas. Las muestras compuestas fueron aclaradas mediante el mismo procedimiento que se usó para las muestras vegetales y se distribuyeron en cinco portaobjetos por temporada y por sitio (20 en total). Para asegurar la homogeneidad de la cantidad de muestra en cada laminilla se empleó una laminilla metálica con orificios de 7 mm de diámetro(21,24). En cada laminilla se identificaron y contabilizaron las estructuras celulares en 20 campos (400 en total) al microscopio. Finalmente, las especies identificadas se clasificaron según su variación temporal (lluvias o secas), forma de crecimiento (arbustiva, arbórea, herbácea, pastos o suculenta) y frecuencia de aparición(25,21). Estos análisis se llevaron a cabo en el Departamento de Zootecnia de la Universidad Autónoma de Baja California Sur.
Análisis de datos
La composición de la dieta se expresó en una matriz donde se incluyeron las frecuencias de cada especie vegetal, de acuerdo con la forma biológica, la época del año y el sitio(26). Además, se evaluó la diversidad de la dieta por sitio y estación del año con el índice de diversidad de Shannon(27). Para conocer si existe diferencia en la diversidad de la dieta por época del año en cada sitio de estudio se aplicó la prueba no paramétrica U de Mann-Whitney (α≤0.05) mediante el software estadístico PAST 4.0. El grado de selectividad del consumo de forraje (ISF) se determinó con base en la disponibilidad y el consumo de cada especie vegetal de acuerdo con el índice de selección de Ivlev(28), mediante la siguiente ecuación:
Ei=ri-p(i)ri+p(i)
Donde; Ei, es el índice de selección de Ivlev; r(i), frecuencia relativa de la especie i en la dieta; y p(i), es frecuencia relativa de la especie en i en el hábitat.
Los valores de selección de Ivlev varían entre -1 (rechazo o selección negativa de una especie) y 1 (preferencia o selección positiva), mientras que un valor de 0 supone un consumo de alimento al azar o en proporción a su disponibilidad. Al respecto, Stuth(29) categoriza los valores de este índice de la siguiente manera: especies con un ISF mayor a 0.35 fueron preferidas sobre otras disponibles; de -0.35 a 0.35, especies de mantenimiento o consumidas de manera proporcional a su disponibilidad; por último, las especies evitadas con un ISF menor a -0.35.
Resultados y discusión
Evaluación de la cobertura vegetal
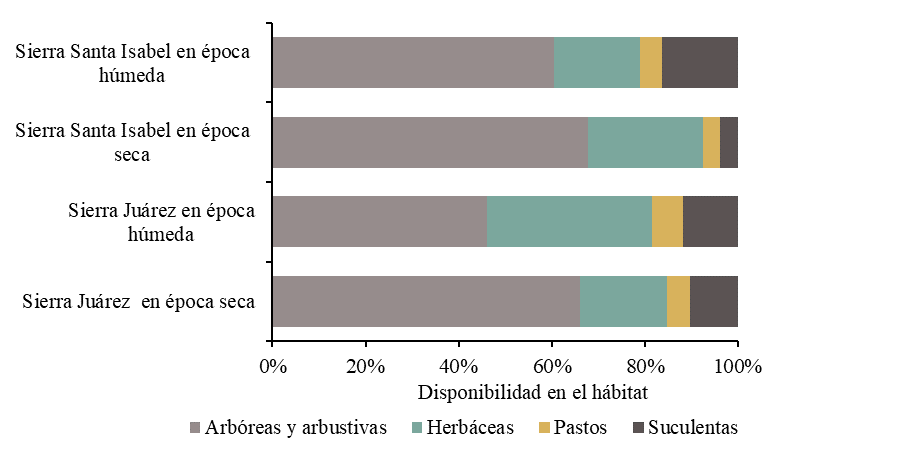
En Sierra Juárez y Santa Isabel, el hábitat del borrego cimarrón se caracterizó por una alta disponibilidad de especies arbóreas y arbustivas, y en menor frecuencia de pastos (Figura 2). La disponibilidad de especies varió en función del sitio y la época del año (P<0.05). En Sierra Juárez se identificaron 52 especies pertenecientes a 23 familias taxonómicas, las principales fueron: Asteraceae (7), Cactaceae (5), Fabaceae (5) y Asparagaceae (4). La diversidad de especies de plantas en esta cordillera fue mayor en la época húmeda (H’= 3.69) que en la época seca (H’= 3.43). En Santa Isabel se identificaron 55 especies correspondientes a 21 familias taxonómicas de las cuales Asteraceae (9), Cactaceae (6), Fabaceae (6) y Euphorbiaceae (5) fueron las más comunes. La diversidad vegetal en Sierra Santa Isabel en la época húmeda (H’= 3.89) fue mayor que en la seca (H’= 3.55). Mantener una diversidad alta de forraje en el hábitat del borrego es importante porque no existe una única especie que cubra todos los requerimientos nutricionales(27).
Figura 2: Variación estacional de la disponibilidad del forraje (%) en relación con el sitio, época del año y forma biológica de crecimiento vegetal
En Sierra Juárez se registraron 52 especies, pero los estimadores de diversidad Chao 2 y Jacknife, predicen una riqueza de 63 especies (82.9 % de efectividad) y 69 especies (75.7 % de efectividad), respectivamente. En Sierra Santa Isabel se identificaron 55 especies vegetales, pero el estimador Chao 2 calcula una riqueza potencial de 66 especies (88.5 % de efectividad), mientras que, con el de Jacknife se estiman 70 especies (78.4 % de efectividad).
Composición y diversidad de la dieta
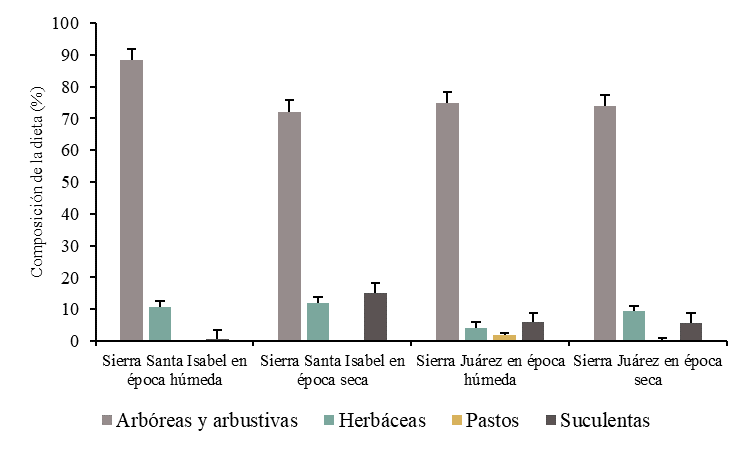
Los herbívoros silvestres están adaptados al consumo de una gran diversidad de tipos de forraje, en función de gradientes ambientales que influyen en la disponibilidad del alimento(30). Al respecto, se registraron 31 especies que conforman la dieta de la población de borrego cimarrón en Sierra Juárez. Durante la época seca se identificaron 25 especies, mientras que, en la época húmeda el consumo ascendió a 27 especies (Cuadro 1). Las formas biológicas de mayor consumo fueron las arbóreas y arbustivas, con una frecuencia de consumo del 74.7 % y 73.8 % en época húmeda y seca, respectivamente. Las herbáceas representaron el 4.11 % de la dieta en la época húmeda y el 9.39 % en la época seca. El aporte de pastos a la dieta fue mayor durante la época húmeda (1.77 %), en tanto que, el de suculentas fue más notable en la época seca (5.64 %; Figura 3). Aunque el índice de diversidad de Shannon mostró valores más elevados durante la época húmeda (H’= 3.05) que en la seca (H’= 2.97), no se encontraron diferencias en la diversidad de la dieta entre ambas épocas del año.
Figura 3: Composición de la dieta del borrego cimarrón en relación con el sitio, época del año y forma biológica del forraje (las líneas verticales sobre las barras indican el error estándar)
La población de borrego cimarrón en Sierra Santa Isabel se alimentó de 43 especies. Durante la época seca la dieta se compuso de 36 especies y en la época húmeda el consumo fue de 29 especies. Las especies arbóreas y arbustivas predominaron en la dieta durante las dos épocas del año: 88.22 % en húmeda y 72.12 % en seca respectivamente. Además, se presentó un 10.76 % herbáceas en época húmeda y 12.12 % en época seca. Se observó una diferencia de 15.15 % (P<0.05) en el consumo de suculentas entre la época húmeda y seca (Figura 3). El índice de diversidad de Shannon mostró valores similares en la composición de la dieta de en la época húmeda (H’= 3.28) y seca (H’= 3.14) esto se debe principalmente a que el mayor porcentaje de consumo corresponde a especies perennes. En ambos sitios de estudio, las especies con mayor frecuencia en la dieta fueron Hibiscus denudatus y L. tridentata (Figura 4).
Figura 4: Especies más frecuentes en la dieta del borrego cimarrón en la Sierra Juárez y Sierra Santa Isabel
A. Gobernadora (Larrea tridentata), de la familia Zygophyllaceae (fotografía de Comisión Nacional Forestal); B. Malva del desierto (Hibiscus denudatus), de la familia Malvaceae (fotografía de James Varnell). Ambas son especies de arbustos perennifoloios nativos de México.
Los estudios sobre los hábitos alimentarios del borrego cimarrón en Norteamérica revelan una alta diversidad de especies en la dieta de estos animales. Monson y Sumner(31) señalan que se han identificado hasta 110 especies de plantas en la dieta del borrego cimarrón en zonas desérticas. En los Estados Unidos de América, específicamente en Arizona(32) reportaron la presencia de 58 especies de plantas en la dieta, mientras que, en California(33) encontraron que los borregos se alimentaron de 32 especies distintas. En regiones del norte de Estados Unidos y del sur de Canadá se ha documentado que la alimentación de los borregos de las Montañas Rocallosas (O. c. canadensis) comprende hasta 200 especies vegetales(33,34).
En Sierra Juárez se registró el menor número de especies en la dieta del borrego cimarrón en comparación con otros estudios realizados en el Desierto Sonorense. En Sonora identificaron 41 especies en la Sierra El Viejo, Caborca(5); en otro estudio(9) encontraron 40 especies en la Sierra Noche Buena, Hermosillo; y O'Farril et al(35) reportaron 39 especies de plantas en la dieta del borrego de la Isla Tiburón. En la Península de Baja California(36) se documentó que la dieta del borrego cimarrón en la Sierra San Pedro Mártir, Ensenada, está compuesta por 72 especies de plantas; se documentó un consumo anual de 47 especies en la Sierra El Mechudo, Baja California Sur(8). En Coahuila, Gastelum-Mendoza et al(6) encontraron un consumo anual de 50 especies. Estos resultados se asemejan a los resultados del presente estudio en cuanto al número de especies consumidas por el borrego cimarrón en la Sierra Santa Isabel(43).
Las variaciones en la composición de la dieta pueden atribuirse a las variaciones fisiográficas y climáticas del hábitat. En este trabajo de investigación se observó que los lugares donde habita el borrego cimarrón son generalmente rocosos, abiertos y con una cobertura vegetal limitada. Este hallazgo, coincide con un estudio sobre la dieta del venado bura en el sur de Arizona(37), donde se observaron altas temperaturas y escasez de lluvias, lo que, influyó directamente en el metabolismo de las plantas, provocando su desecación en poco tiempo.
La intensidad de herbivoría sobre algunas especies puede repercutir negativamente en la dinámica de la vegetación. Al respecto, las especies identificadas en la dieta del borrego cimarrón se clasificaron en decrecientes o básicas (las de mayor consumo que disminuyen su disponibilidad en función de la alta presión de herbivoría) y crecientes o de emergencia (las de menor consumo que aumentan su disponibilidad en función de la baja presión de herbivoría)(30,38).
De acuerdo con la composición de la dieta del borrego cimarrón en Sierra Juárez se clasificaron como especies básicas H. denudatus y L. tridentata, que, en conjunto, aportaron el 21.82 % de la dieta anual. Sin embargo, estas especies solo representaron el 6.5 % de la riqueza total de especies que conforman la dieta. Asimismo, se consideraron como especies de emergencia 11 que en conjunto aportaron el 9.87 % de la dieta y el 35.5 % de la riqueza de especies consumidas. En la población de Sierra Santa Isabel, se consideraron como especies básicas H. denudatus, L. tridentata, Solanum hindsianum, Condea emoryi y Eriogonum inflatum, las cuales representaron en conjunto el 40.48 % de la dieta anual, pero solo el 11.63 % de la riqueza total de especies que conforman la dieta. Por el contrario, 15 especies se consideraron crecientes, ya que en conjunto aportaron el 9.8 % de la dieta anual, y 13 de ellas, aportaron cada una menos del 1 % de la dieta. Estas especies pueden considerarse de emergencia y son importantes en periodos de baja disponibilidad de especies básicas(6). Por ejemplo, Amaranthus palmeri, Atriplex barclayana, Senegalia greggii y Krameria erecta (Cuadro 1).
Las especies arbóreas y arbustivas constituyeron la base de la dieta del borrego cimarrón (Figura 3). Este resultado coincide con otros estudios sobre los hábitos alimentarios de esta especie en Norteamérica(8,9,35). La importancia de las especies arbustivas y arbóreas en la dieta del borrego se debe a que son forrajes disponibles durante todo el año(6). Asimismo, Bolen y Robinson(39) afirman que los herbívoros silvestres en zonas áridas prefieren el ramoneo de arbustos y árboles porque contienen más nutrientes digestibles que otras especies de pastos. De acuerdo con algunos investigadores(40) los arbustos en zonas áridas acumulan reservas de nutrientes durante su crecimiento para la formación de nuevos tejidos, lo que resulta en una mayor concentración de proteína cruda en comparación con algunas hierbas y pastos.
La selección del forraje varió en función del sitio y la época del año (P<0.05). En Sierra Juárez durante la época seca, el borrego cimarrón prefirió el consumo de H. denudatus, L. tridentata y E. inflatum; mientras que, en la época húmeda tuvo preferencia por Bebbia juncea y Justicia californica. En la época seca evitó consumir Cylindropuntia ramosissima, Ephedra californica y Sphaeralcea ambigua; y durante la época húmeda las especies que evitó fueron Neltuma glandulosa y Krameria bicolor (Cuadro 2). Algunas de estas especies son evitadas por presentar estructuras que impiden su consumo óptimo, por ejemplo, espinas, pubescencias o altos contenidos de cera en sus hojas y tallos(30). Cabe destacar que se identificaron especies poco comunes en la dieta del borrego como Washingtonia sp. y Typha domingensis (Cuadro 1) las cuales crecen en sustratos húmedos alrededor de las fuentes de agua naturales.
Se identificó una diferencia significativa (P<0.05) entre la disponibilidad y el consumo de especies herbáceas. Aunque éstas presentaron una alta disponibilidad, particularmente en la Sierra Juárez (Figura 2), no se observaron en porcentajes elevados de consumo (Figura 4). Las herbáceas, especialmente las especies anuales, tienden a ser menos consumidas por los herbívoros silvestres en zonas áridas(5,6,9), dado que su disponibilidad está estrechamente vinculada a la humedad y las precipitaciones. Como resultado, su presencia en la cobertura vegetal se limita a breves periodos del año. Aunque estudios previos indican que las herbáceas son más comunes en la dieta de los herbívoros durante la temporada húmeda(6,30), en el presente estudio se detectaron los mayores porcentajes de aparición en las dietas durante la época seca. Esto puede explicarse por la existencia de oasis en las áreas de estudio, que proporcionan humedad suficiente para el crecimiento de las hierbas durante todo el año. A pesar de que su contribución a la dieta general fue baja, las herbáceas desempeñan un papel crucial en la nutrición del borrego cimarrón. Estas plantas ofrecen entre un 35 y un 40 % más de energía, niveles de proteína similares y un 40 a 45 % más de fósforo en comparación con los arbustos del norte de México(12). Además, son particularmente relevantes durante la temporada reproductiva. Gastelum-Mendoza et al(9) reportaron que tanto machos como hembras de borrego cimarrón consumían principalmente herbáceas durante este periodo en la Sierra Noche Buena, Sonora, con un consumo del 38.6 y 47.6 %, respectivamente. Finalmente, aunque las herbáceas no fueron altamente consumidas en general, se observó una preferencia significativa por E. inflatum en la Sierra Santa Isabel a lo largo de todo el año (Cuadro 3).
No obstante que la fisiología digestiva del borrego cimarrón está adaptada para la digestión de pastos(41), los cuales suelen presentar altos contenidos de fibra y una digestibilidad relativamente baja durante la mayor parte del año(8,30), estos no fueron componentes significativos en la composición de su dieta (Figura 3). Al respecto, Brown et al(34) señalan que las poblaciones de borregos silvestres de montaña en Nevada, EUA, consumen entre el 62 a 81% de pastos comparado con las poblaciones de borregos de desierto. Esto debido a una mayor disponibilidad de arbustos y árboles en el Desierto Sonorense(42). Lo anterior, coincide con hallazgos previos en la Sierra El Mechudo, Baja California Sur, donde solo se identificaron dos especies de pastos en la dieta(8). De manera similar, en el estado de Sonora, estudios realizados por Tarango et al.(5) en la Sierra El Viejo y por O'Farrill et al.(35) en la Isla Tiburón reportaron que los pastos representaron solo un 5 % de la dieta. Sin embargo, investigaciones más recientes encontraron un aporte de pastos del 26.8 % en la Sierra Noche Buena, Sonora(9) y 17.21 % en un matorral desértico rosetófilo en Coahuila(6). También se señala(30) que en las zonas áridas del norte de México los pastos tienen un alto contenido de celulosa y hemicelulosa, lo que limita su valor nutricional y reduce su consumo por parte de los herbívoros silvestres. En este sentido, se menciona(12) que Aristida adscensionis presenta valores de fibra detergente neutra (compuesta por celulosa, hemicelulosa y lignina) del 61.3 % en 2010 y del 79.6 % en 2011 en la dieta del borrego de la Sierra el Mechudo, Baja California Sur. Estos valores sugieren que se trata de una especie de pasto con digestibilidad baja(43).
En Sierra Santa Isabel, el aporte de especies suculentas a la dieta del borrego cimarrón fue mayor que en Sierra Juárez (Figura 3) y varió en función de la época del año (P<0.05); con mayor consumo de estas plantas en la época seca. Las especies suculentas no representan un aporte nutritivo alto para el borrego(12), pero son elementos importantes en la dieta como una fuente importante de agua en la época seca(5,6,30) cuando requiere un mínimo de consumo de agua equivalente al 4 o 5 % de su peso corporal(42). La relevancia de los nopales, magueyes y biznagas en la dieta del borrego se ha reportado en diferentes estudios: en Sonora, en la Sierra El Viejo, la frecuencia del consumo de suculentas fue del 18 %(5,9); en la Sierra El Mechudo, fue del 0.2 %(8); en la Sierra San Pedro Mártir, Baja California fue del 12 %(44).
Conclusiones e implicaciones
En Sierra Juárez y Sierra Santa Isabel, el borrego cimarrón se alimentó principalmente de especies arbóreas y arbustivas. Mientras que, las suculentas fueron especies importantes en la dieta durante la época seca. Aunque no se detectaron diferencias significativas en la diversidad de la dieta entre ambas sierras, las especies clave en la dieta del borrego cimarrón fueron diferentes entre sitios. En Sierra Juárez, las principales especies consumidas fueron E. inflatum, H. denudatus, J. californica y L. tridentata; mientras que, en Sierra Santa Isabel fueron E. inflatum, H. denudatus, Horsfordia newberryi y L. tridentata. Estos resultados son importantes para identificar y delimitar áreas prioritarias de forrajeo para las poblaciones de borrego cimarrón, así como para guiar el diseño de estrategias comunitarias de manejo de especies forrajeras en el estado de Baja California. Además, se recomienda complementar estos hallazgos con un análisis bromatológico de las principales especies consumidas para evaluar su calidad nutricional y su impacto en la salud de las poblaciones de borrego cimarrón.
Agradecimientos
Los autores agradecen a la Universidad Autónoma de Baja California por el financiamiento de esta investigación a través del proyecto 400/2975 y a la Fundación UABC por gestionar el financiamiento de la misma.
Conflicto de intereses
Los autores declaran no tener conflicto de intereses.
Cuadro 1: Composición de la dieta del borrego cimarrón de acuerdo con la frecuencia relativa de las especies identificadas en las muestras fecales, sitio de recolecta y época del año
Cuadro 2: Tipos de uso y selección del forraje por el borrego cimarrón de acuerdo con el índice de Ivlev, sitio y época del año
Tipo de uso: proporcional (P), seleccionada (S) y evitada (E).
Literatura citada: