https://doi.org/10.22319/rmcp.v16i1.6669
Nota de investigación
Análisis preliminar del desarrollo de un programa de crianza del Caballo Peruano de Paso en condiciones de campo
Jorge Luis Vilela Velarde a*
Paola Giovanna María Quintana Dolores a
María Lourdes Velarde Marcos a
Teresa Vargas Rodríguez a
Rosario Gonzales Lecaros a
José Dextre Chacón a
a Universidad Científica del Sur. Facultad de Ciencias Veterinarias y Biológicas. Carrera de Medicina Veterinaria y Zootecnia. Grupo de Investigación en Ciencia Equina y Animales Autóctonos. Lima, Perú.
*Autor de correspondencia: jvilela@cientifica.edu.pe
Resumen:
Hasta la fecha no se han determinado los parámetros genéticos de sobrealcance, término y agudez en Caballos Peruanos de Paso (CPP). Es importante estimar estos parámetros para su aplicación en la crianza de CPP, por lo tanto, el objetivo de este estudio fue estimar la heredabilidad, repetibilidad y correlaciones genéticas en condiciones de campo del sobrealcance, término y agudez de CPP. El estudio incluyó 134, 137 y 134 registros de sementales y yeguas para los rasgos de sobrealcance, término y agudez, respectivamente. Todas las mediciones se grabaron en formato de vídeo MP4 con una resolución de 1,920 x 1,080 megapíxeles y a 60 fotogramas por segundo. Todos los rasgos se midieron de tres a cinco veces (una vez por zancada) y se analizó cada rasgo. Para el análisis de las mediciones se utilizó el software KINOVEA versión 0.9.5. Se utilizó un modelo animal multivariado de medidas repetidas con efecto del sexo para estimar los componentes de la varianza para cada rasgo utilizando el software WOMBAT. Los resultados mostraron una heredabilidad de 0.411, 0.476 y 0.405, para los rasgos de sobrealcance, término y agudez, respectivamente. La repetibilidad fue alta en todos los rasgos (> 0.70). Las correlaciones genéticas aditivas oscilaron entre -0.30 y 0.49. Se puede concluir que el sobrealcance y el término tienen altos valores de heredabilidad, lo que permite que estos rasgos respondan mejor en un proceso de selección, a diferencia de la agudez, que tiene un valor de heredabilidad moderado.
Palabras clave: Heredabilidad, Correlaciones genéticas, Repetibilidad, Sobrealcance, Término, Agudez.
Recibido: 03/04/2024
Aceptado: 23/10/2024
El Caballo Peruano de Paso (CPP) es una raza equina originaria de Perú. Desde 1947 la Asociación Nacional de Criadores y Propietarios de Caballos Peruanos de Paso (ANCPCPP) de Perú ha realizado grandes esfuerzos en la conservación, crianza y selección de esta raza(1). Esta raza es considerada un caballo de paso con un ritmo simétrico de cuatro tiempos y una secuencia de pisadas laterales durante el andar de paso llano(2-4). La ANCPCPP(5) define el paso llano como cuando el caballo rompe la ambladura por los lados en 4 pasos. Otras razas tienen andares similares, aunque con algunas diferencias, incluyendo el clásico Fino, el rack ondulado, el rack de mapache, el trote de zorro, la marcha picada, el rack de placer de montaña, el rack de montaña rocosa, el andar de camino, el sobreandando y el toelt(4). En CPP, la suavidad y armonía del movimiento surge de la combinación de modalidades de ejecución durante el paso llano (o ambladura rota)(2). Los principales rasgos implicados son el término, la agudez(6-9) y el sobrealcance(10-13) (Figura 1). Se desean valores óptimos de estos tres rasgos utilizados como parte de los criterios para evaluar el desempeño de cada animal. Estos rasgos fueron seleccionados porque influyen directamente en la suavidad, armonía y eficiencia del movimiento del caballo, resultando en un andar suave(7), que es la característica que distingue a esta raza de otras en el mundo, y es de suma importancia en la selección de caballos por parte de los criadores. La evaluación de estas características durante el movimiento permite identificar a los animales con los movimientos más favorables y, por lo tanto, con mayor potencial para ser utilizados en programas de mejoramiento genético. Al evaluar caballos, la medición de los rasgos por el ojo humano puede ser un desafío, debido a la subjetividad y a la precisión limitada(9,14). Varios estudios sobre medicina deportiva equina se han centrado en la cinemática en caballos, identificando cambios en el rendimiento deportivo y la salud(8,15,16). Para medir los rasgos funcionales de un caballo, se recomienda estimar variables cinemáticas objetivamente medibles(17). Por lo tanto, el objetivo de este estudio fue estimar la heredabilidad, repetibilidad y correlaciones genéticas en condiciones de campo del sobrealcance, término y agudez en el CPP.
Para lograr los objetivos propuestos, se fenotiparon 140 animales, de los cuales solo se utilizaron grabaciones que pudieron ser analizadas. Las grabaciones no consideradas en el análisis se descartaron debido a problemas de grabación relacionados con la lente de la videograbadora. Los caballos que no se movían paralelos a la cámara, que cojeaban, tenían una velocidad muy lenta, el adiestrador bloqueaba la vista de la cámara o yeguas con potros a los lados durante la marcha que impedían la identificación de las marcas durante el paso llano, no fueron incluidos en el estudio después de depurar los videos. Finalmente, se utilizaron los registros de 134, 137 y 134 animales, con una mayor proporción de hembras que de machos (80 % y 20 %, aproximadamente), para estudiar los rasgos de sobrealcance, término y agudez, respectivamente. La edad media de los caballos fue de 7.67 ± 2.61 años, oscilando entre los 5 y los 11 años (mediana de 7.05 años). Se utilizaron los mismos animales para el análisis de los tres rasgos. De acuerdo con cada rasgo (sobrealcance, término y agudez), se incluyeron en la base de datos un total de 1,615, 1,641 y 1,615 individuos, respectivamente, que podían rastrearse 21 generaciones atrás. En el Cuadro 1 se presenta información relacionada con el análisis de pedigrí, incluido el número de animales rastreados, los fenotipos relacionados y los coeficientes de consanguinidad. El intervalo generacional de toda la población rastreada fue de 8.76 ± 4.53 años. Este estudio fue aprobado por el Comité Institucional de Ética en Investigación con Animales y Biodiversidad de la Universidad Científica del Sur (Cod. 028-2021-PRO99) y se obtuvo permiso de los propietarios de los animales para la recolección de datos. Solo se incluyeron en el estudio animales sanos, examinados por un veterinario registrado, sin signos de cojera en una o más patas.
Cuadro 1: Estructura del pedigrí para cada rasgo
Todos los animales fueron evaluados en sus instalaciones de crianza. Estos consistían en campos cubiertos de pasto, planos, secos y sin obstáculos. Se determinaron los puntos de inicio y finalización, los cuales fueron perpendiculares a la ubicación de la lente de la cámara, a través de los cuales se movió cada animal durante la grabación de sobrealcance y agudez. Durante la grabación, cada caballo recorrió una distancia recta de 50 m desde un punto de salida hasta un punto de llegada ubicado frente a la lente de la cámara. Cada caballo fue evaluado en diferentes días, con grupos de tres a diez caballos evaluados por día, dependiendo de la disponibilidad de los criadores. Cada criador utilizó un adiestrador experimentado para grabar los videos.
Para identificar los puntos de referencia para medir cada rasgo, se colocó una cinta de 4 x 4 cm en las áreas marcadas en la Figura 1. Todas las mediciones se grabaron en formato de vídeo MP4 con una resolución de 1,920 X 1,080 megapíxeles y a 60 fotogramas/seg(18). Los animales se colocaron sobre una superficie plana y se jalaron por un operario a paso llano con una velocidad aproximada de entre 2.5 y 4 m/seg, cubriendo 50 m. La actuación del caballo se grabó filmando con cámara de video en un trípode con una posición fija colocada horizontalmente (confirmada con un nivel) a una altura de 1.3 m y 12 m desde el centro de la línea de movimiento, grabando el movimiento de cada animal lateralmente. Todos los rasgos se midieron mediante tres a cinco repeticiones técnicas (una por zancada) y se incluyeron en el modelo para su análisis. La descripción de la medición de cada rasgo se detalla en la Figura 1. Para el análisis de las mediciones se utilizó el software KINOVEA versión 0.9.5 (http://www.kinovea.org/).
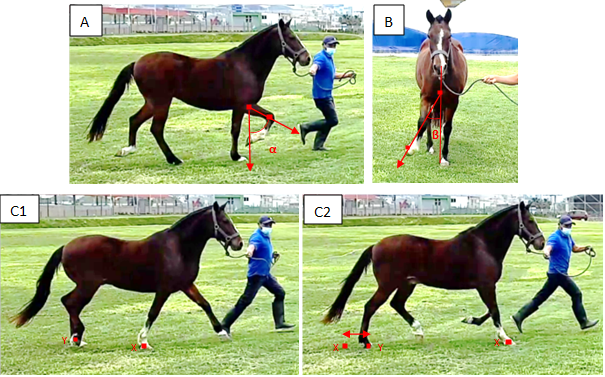
Figura 1: Parámetros genéticos de sobrealcance, término y agudez en Caballos Peruanos de Paso
Agudez (A): representación del ángulo α máximo de la agudez de la extremidad anterior derecha medido desde la orientación que conecta la rodilla con el codo con respecto a la vertical, en el plano sagital (9). Término (B): representación del ángulo β máximo del término de la extremidad anterior derecha medido desde la pared lateral del casco con respecto a la vertical en el plano coronal del caballo (9). Sobrealcance (C1 y C2): representación del sobrealcance, medido como la distancia entre X e Y, donde X es la pisada de la posición del casco de la extremidad anterior derecha durante la máxima protracción y retracción en el suelo e Y es la pisada de la posición del casco de la extremidad trasera derecha durante la máxima protracción y retracción en el suelo cuando se excede X en la siguiente zancada.
Para el análisis estadístico, todos los rasgos se sometieron a análisis estadístico descriptivo y análisis de normalidad mediante la prueba de Anderson-Darling con P>0.05 indicando que el rasgo cumplió con la distribución normal. Para estos análisis se utilizó el software JASP. La heredabilidad se expresa por h2=a2p2, donde a2 es la varianza genética aditiva, y p2 es la varianza fenotípica(20). Los registros fenotípicos de los tres rasgos se ajustaron al modelo animal multivariado de medidas repetidas con efecto fijo del sexo para estimar los componentes de la varianza de cada uno de los tres rasgos utilizando el algoritmo de información promedio (IP) para máxima verosimilitud restringida(21). Para todos los procedimientos se utilizó el software WOMBAT (http://didgeridoo.une.edu.au/km/wombat.php)(22). El modelo utilizado se expresa como sigue:
Yijk=μ+Sexoi+Animalj+Caballok+ eijk,
Donde:
Yij es el valor fenotípico de cada rasgo;
μ es la media poblacional;
Sexoi es el efecto fijo del sexo (2 niveles);
Animalj es el efecto aleatorio del j-ésimo animal ~ND0,Aa2, A, denota la matriz de relaciones del numerador entre los animales y a2 la varianza aditiva;
Caballok es el efecto aleatorio de la k-ésima medida del individuo (3 a 5 niveles) ~ND0,Ipe2, donde I es la matriz de identidad, pe2 es la varianza permanente del ambiente;
eijk es el efecto aleatorio residual ~ND0,Ie2.
Se utilizó la estructura de pedigrí para la estimación de varianzas aditivas y covarianzas del efecto aleatorio para el cálculo de heredabilidad, repetibilidad y correlaciones genéticas.
La repetibilidad puede estimarse como la proporción de la varianza fenotípica explicada por la varianza aditiva y la varianza ambiental permanente(20). Las contribuciones no aditivas a las diferencias consistentes entre individuos se denominan normalmente “efectos ambientales permanentes”. Si un rasgo tiene medidas repetidas, entonces es necesario modelar los efectos ambientales permanentes en un modelo animal para prevenir el sesgo al alza en la varianza aditiva(22). Por lo tanto, la varianza entre individuos está dada por el componente Animalj, mientras que el componente residual representa la varianza dentro del individuo(22). Para el cálculo de la repetibilidad, se utilizaron tres registros (3 niveles) por animal por rasgo para estimar estos componentes de la varianza. La repetibilidad (R) se calculó a partir de: R= a2+ pe2a2+ pe2+ e2 . Las correlaciones genéticas fenotípicas y aditivas se calcularon con los mismos registros utilizados para los cálculos de heredabilidad. Todos los rasgos se sometieron a la prueba de normalidad de Anderson-Darling y mostraron una distribución normal con un valor de significancia de P>0.05 para todos los rasgos, por lo tanto, el uso del modelo animal propuesto es apropiado.
De acuerdo con los resultados, se encontró que el sobrealcance tiene una alta(23) heredabilidad, en línea con los hallazgos de Molina et al(24) para la longitud de zancada. En contraste, Sole et al(10) reportaron una heredabilidad de sobrealcance en el caballo Lusitano que fue considerablemente menor que el del presente trabajo. Además, se observó que el sobrealcance en el presente estudio difiere significativamente del presentado en otros estudios, particularmente en la marcha de trote(10). Sin embargo, también se reportaron(12,13) sobrealcances positivos en caballos andaluces. La heredabilidad del término también resultó ser alta. En cuanto a la agudez, su alta heredabilidad fue similar a la encontrada en los caballos Lusitanos(10), aunque por un rasgo relacionado, pero diferente. La discrepancia en los resultados en comparación con Molina et al(24) podría ser atribuida a factores como el nivel de entrenamiento del caballo(25) y la omisión del efecto del sexo en su estudio, ya que solo evaluaron a los machos.
Como se pudo observar, los valores de heredabilidad de todos los caracteres analizados fueron altos (mayores a 0.40). Estos valores pueden ser explicados por la no inclusión de un factor externo que permita el andar libre del caballo, como un jinete que pueda alterar el ritmo y el movimiento del animal durante la marcha plana(26). Estos movimientos se realizan libremente y son más homogéneos sin el efecto externo y, por lo tanto, son más heredables(27). Otra razón que podría explicar los altos valores de heredabilidad es que puede haber habido una mayor especialización en los concursos de CPP(27), o un mejor uso del proceso de selección en la raza(23), aunque también puede deberse a que esa población fue más homogénea debido al mayor número de yeguas (~80 %) analizadas en este estudio y solo se incluyeron animales adultos (de 5 a 11 años)(27). Una forma de lograr un mayor progreso genético puede ser con una mayor intensidad de selección, así como con mayores valores de heredabilidad(28). Además, también debe tenerse en cuenta que la mejora de estos rasgos es el resultado de una compleja combinación de rasgos conformacionales, fisiológicos y conductuales(29). La eficiencia en la selección genética de variables biocinemáticas puede ser más eficiente que la selección basada en el desempeño de los animales, y eso puede traducirse en mayores valores de heredabilidad(30).
Como criterio para categorizar las correlaciones, se utilizó la escala de la Universidad de Quinnipiac(31) para clasificar las correlaciones menores o iguales a 0.20 como débiles, mayores que 20 y menores que 0.40 como moderadas y mayores que 0.40 como fuertes(27,32,33). Las correlaciones genéticas aditivas encontradas en el presente estudio se clasificaron entre valores absolutos de 0.301 (error estándar= 0.432) y 0.697 (error estándar= 0.374), semejantes a otros estudios realizados en condiciones de campo(30), y las correlaciones fenotípicas se clasificaron entre valores absolutos de 0.183 (error estándar= 0.081) y 0.213 (error estándar= 0.079). Si bien es cierto que las correlaciones genéticas proporcionan información sobre la relación entre los rasgos, no siempre son tan útiles como las correlaciones fenotípicas en el momento de la evaluación durante el desempeño(32), posiblemente debido al tiempo de entrenamiento en el momento de la evaluación, la experiencia del jinete u otros factores ambientales. Su principal utilidad puede ser aplicada a la construcción de índices de selección o para predecir la respuesta correlacionada a la selección(34).
En cuanto a los valores de repetibilidad, estos fueron superiores a 0.70 para todos los rasgos, lo que se considera excelente(35). Se encontraron resultados similares en los rasgos cinemáticos en caballos entrenados de doma clásica y taurinos (más de 0.50), y en caballos suecos de sangre caliente en marchas puntuadas con valores entre 0.75 y 0.77(36). Los bajos valores de error estándar cercanos a 0.02 indican alta precisión en todos los rasgos (0.898, 0.842 y 0.901 para sobrealcance, término y agudez, respectivamente), lo que indica que este parámetro tiene poco efecto en el ambiente temporal de los tres rasgos(20). Esto también puede ser corroborado por los valores de c2 obtenidos en este estudio, con valores que van de moderado a alto, lo que indica que una proporción considerable de la varianza fenotípica es explicada por la varianza aditiva y la varianza ambiental permanente, más que por la varianza ambiental temporal(20), aunque esto no es conciso con el rasgo de término, que puede ser afectado principalmente por el efecto genético aditivo. Teniendo en cuenta la metodología de Sepúlveda et al(35), quienes encontraron mayores valores de repetibilidad en las observaciones diarias que en las semanales, se puede sugerir que las observaciones realizadas con diferencias mínimas podrían ser aún mayores, tal como se encontró en este estudio. Esto puede ser corroborado en un estudio realizado en caballos de diferentes razas(37) en el que la repetibilidad de la asimetría de la posición de la cabeza y la pelvis presentó valores entre 0.89 y 0.95. Además, los resultados del presente estudio corroboran que los rasgos con alta repetibilidad requieren pocas mediciones (3 en este estudio) para obtener una mayor precisión, y un aumento en el número de mediciones puede ser irrelevante para la estimación de parámetros. Teniendo en cuenta los altos valores de repetibilidad encontrados, este parámetro puede ser utilizado como indicador de cuán efectivo puede ser el proceso de selección, considerando su relación con la heredabilidad, debido a la inclusión de la varianza ambiental permanente (varianza intraindividual) en su estimación(20,38).
Esta investigación presenta varios hallazgos positivos, con valores de heredabilidad prometedores que indican que el sobrealcance, el término y la agudez tienen un alto potencial de mejora en un plan de selección. Las correlaciones genéticas positivas encontradas entre el sobrealcance y la agudez sugieren que ambos rasgos pueden mejorarse simultáneamente a través de estrategias de mejoramiento específicas. Además, los altos valores de repetibilidad con alta precisión indican que se puede reducir el número de mediciones necesarias para estos rasgos, lo que simplifica el proceso de evaluación.
Sin embargo, existen ciertas limitaciones en el estudio. La precisión general de los rasgos fue algo limitada, probablemente debido al pequeño tamaño de la muestra. La complejidad de realizar las mediciones cinemáticas y el tiempo requerido para viajar entre los diferentes centros de crianza fueron las principales razones de esta limitación. Sin embargo, a pesar del pequeño número de animales fenotipados, la estimación de los parámetros genéticos se justifica como valores preliminares y son útiles para referencias posteriores, como se ha observado en otros estudios con tamaños de muestra similares (100 a 362)(29). La población de animales incluidos en este estudio fue pequeña, pero el coeficiente de consanguinidad total promedio (~5.43 %) fue comparable a los estudios de Larrea et al(39) y Montenegro et al(40) (5.97 % y 5.44 %, respectivamente). Esto sugiere que los resultados obtenidos pueden interpretarse como una referencia para la población general de CPP. Todos los caballos fueron evaluados con el mismo dispositivo en condiciones de campo, asegurando que cualquier sesgo potencial fuera consistente entre los sujetos.
En conclusión, los hallazgos de este estudio proporcionan una referencia valiosa para el mejoramiento genético de la CPP, a pesar de las limitaciones señaladas. Los resultados indican que los rasgos, como el sobrealcance, el término y la agudez, exhiben una alta heredabilidad y pueden ser abordados de manera efectiva en los programas de mejoramiento. Los parámetros genéticos preliminares del estudio y la comparabilidad de los coeficientes de consanguinidad con otras investigaciones respaldan la relevancia y aplicabilidad de estos resultados.
Conflictos de intereses
Los autores afirman que se utilizaron registros de ocho animales propiedad de José Dexte. También se señala que José Dextre era Presidente del Consejo de la Universidad Científica del Sur durante el desarrollo de la fase metodológica de esta investigación.
Anexo 1: Estadísticos descriptivos por rasgo
IC= intervalo de confianza.
Anexo 2: Estimaciones de la heredabilidad (h2), relación entre la varianza ambiental permanente y la varianza fenotípica (c2) y la repetibilidad (R) (sobre la diagonal), correlaciones fenotípicas (debajo de la diagonal) y correlaciones genéticas aditivas (por encima de la diagonal)
Error estándar entre paréntesis.
Literatura citada: