https://doi.org/10.22319/rmcp.v16i1.6654
Review
Control and bacterial dissemination associated to cell death in Mycobacterium bovis infection. Review
Paola Andrea Ortega-Portilla a
Omar Escobar-Chavarría b
José Ángel Gutiérrez-Pabello a*
a Universidad Nacional Autónoma de México. Facultad de Medicina Veterinaria y Zootecnia. Departamento de Microbiologia e Inmunologia. Laboratorio de Investigación en Tuberculosis y Brucelosis. Ciudad de México, México.
b Universidad Nacional Autónoma de México. Facultad de Estudios Superiores Cuautitlán. Estado de México, México.
*Corresponding author: jagp@unam.mx
Abstract:
One of the hallmarks of Mycobacterium bovis infection is cell death. The type of cell death occurring during the infection determines the persistence of mycobacterial diseases. The aim of this article is to provide a comprehensive review and draw the possible scenarios of cell death types in the pathogenesis of bovine tuberculosis. The current data suggest that: 1) the development of apoptosis and its different variants is related to mycobacterial control, 2) autophagy is a conserved mechanism that limits mycobacterium intracellular replication, 3) pyroptosis is an extreme mechanism that helps control M. bovis at the cost of damaging host tissue, and 4) necrosis will allow the escape and proliferation of mycobacteria.
Keywords: Cell death, Bovine tuberculosis, Mycobacterium bovis, Apoptosis, Autophagy, Pyroptosis.
Received: 06/03/2024
Accepted: 30/09/2024
Introduction
The way a cell dies plays a crucial role in physiological processes. In mycobacterial infections, some types of cell death have been cataloged as defense mechanisms of the host but also as consequences of the pathogen's virulence factors(1,2,3). The Nomenclature Committee on Cell Death (NCCD) 2018, proposes classifying the types of cell death based on the mechanistic and essential aspects of the process, categorizing the majority within the group of regulated cell death(4). Of this large group, some types have been reported in mycobacterial infections, for example, apoptosis(5-9), pyroptosis(10), ferroptosis(11), and necroptosis(12).
Mycobacterium bovis (M. bovis) belongs to the Mycobacterium tuberculosis complex. This species is the causative agent of zoonotic tuberculosis and the main etiologic agent of bovine tuberculosis (bTB). M. bovis affects many animal species(13,14); therefore, it is a problem for public health and the livestock sector(15,16).
M. bovis is mainly transmitted by air, through exhaled droplets from the respiratory system of infected animals. A cellular immune response is developed, which is considered the main immune mechanism against intracellular bacteria(17,18). The dinamics between, macrophages, neutrophils, fibroblast, dendritics cells, B cells, γδ T cells, CD4+, CD8+ lymphocytes, and pro-inflammatory cytokines such as tumor necrosis factor (TNF) and interferon gamma (IFN-γ), give rise to the formation of the characteristic defense structure against mycobacteria: the granuloma(19,20,21).
Granulomatous lesions are characteristic of bTB, and are found mainly in lymph nodes and the lungs(22). Its development varies in different lymph nodes of the same animal(23) and in addition, there has been reported variation in the structural morphology of granulomas from calves and adults(24), therefore granulomas in bTB are considered to have a heterogeneous presentation.
Cell death is one of the determining mechanisms in the formation and evolution of the granuloma that drives the development of the infection and presentation of the disease(25). As a consequence of the persistent nature of M. bovis infection, several types of cell death may occur, highlighting apoptosis and necrosis(5-9,25). However, other modalities, such as pyroptosis and autophagy, may also play a role in the infection(10,26,27,28). This review aims to provide a comprehensive summary of the types of cell death that have been identified in M. bovis infection and highlight their impact on the host. To achieve this goal, we present the information divided in two main sections: 1) Pathogenesis and immune response in bTB and 2) cell death pathways in bTB.
Pathogenesis and immune response in bovine tuberculosis
Bovine tuberculosis is transmitted by direct contact with infected excretion products (urine, saliva, milk, semen, uterine discharges) or mycobacteria present in exhaled droplets from the respiratory system of infected animals(29). The respiratory system is mainly affected, including the lungs and associated lymph nodes(24,30,31). Lesions in the digestive system have been related with transmission by ingestion of contaminated food(32), and transplacental transmission occurs in calves born with congenital infection(33).
bTB can be subclinical for long periods, symptomatic (fever, weight loss, respiratory distress, and decreased milk production), or have an evolution towards a generalized presentation as a consequence of the lymphatic or hematogenous dissemination of the mycobacteria changing to the early and late phases of the infection(14). Factors such as the localization of the disease, the evolution of the primary lesion, mycobacterial virulence factor, bacterial concentration, development of granulomatous lesions and immunocompetence of the host, are determinants for the presentation of clinical symptoms(15,21,29,34,35).
The immune response plays a crucial role in the evolution of the infection in acute and chronic phases(36). In particular, the cell-mediated responses are vital(37). Since the respiratory system is one of the most affected by M. bovis, transcriptional and functional studies have been carried out in different cell populations of this system.
Alveolar macrophages are among the first cell populations infected by inhaled mycobacteria; therefore, they have been studied using different approaches. Transcriptomic analyses have revealed that the changes in gene expression are contrasting. For instance, a decrease in the expression of genes relevant to the recognition of M. bovis(37,38), and a greater polarization of macrophages towards a more permissive-replicative M2 phenotype(39) have been reported. On the other hand, genes that encode chemokines, recognition receptors and proinflammatory molecules showed an increase upon infection with M. bovis(40,41). And finally, this approach has also identified genomic variation related to both susceptibility/resistance to infection(42). Another study using composition and lipid metabolism analysis, identified significant differences in the lipid group between M. tuberculosis related to the formation of foamy macrophages and M. bovis with the inhibition of autophagy(43).
These findings related to the protective and non-protective function of alveolar macrophages against infection, accompanied by a response dependent on mycobacterial species, demonstrate the determining role played by the mycobacteria/alveolar macrophage interaction both in the acute phase and in the evolution of the infection.
Neutrophils are another cell population important in mycobacterial infection(44). Bovine neutrophils function as regulatory cells mainly in the innate immunity of clinical healthy cattle, but also in infected conditions(45). For instance, bovine neutrophils exposed to M. bovis increased phagocytosis, cellular activation, secretion of pro-inflammatory cytokines and intracellular replication(46). These results suggest that M. bovis infection could modulate the response in bovine neutrophils.
β-defensin-5 is an antimicrobial peptide stored in bovine macrophages and neutrophil granules. Incubating recombinant β-defensin-5 with M. bovis evidenced its time-dependent antimicrobial effects; this peptide inhibited growth by 88 % and disrupted the mycobacterial wall at 72 h of incubation(47). The immunoprotective role of recombinant β-defensin-5 was also demonstrated. Recombinant β-defensin-5 from bovine neutrophils was used in the immunization of mice, which were then infected with M. bovis. The results showed a reduction in inflammatory tissue and in the bacterial load in the lung and spleen, demonstrating the potential of its immunoprotective function(48).
The changes in the structure of neutrophil nuclei have been suggested as a complementary diagnostic method for bovine tuberculosis. In human neutrophils exposed to serum from Purified Protein Derivative from Mycobacterium (PPD+) cows, after 3 h, pyknocytosis was the most common change observed in cell nuclei(49). Additionally, specific pattern of expression of IFN-inducible transcriptional genes, myeloperoxidase(MPO) and pentraxin-related protein pentraxin-inducible protein (PTX3) genes, from neutrophils showed their potential as diagnostic tools for M. bovis infection in cattle(50). Despite the modulatory effect that M. bovis apparently exerts on neutrophils, the antimicrobial findings of some of its intracellular molecules evaluated in a recombinant manner, could represent a field of research for biotechnological development with the potential for application in diagnosis and therapeutics.
Considering the importance of dendritic cells (DCs) in innate and adaptive immunity, some research groups have studied their response against M. bovis infection. A comparative analysis between bovine DCs and macrophages, both infected with M. bovis, identified lower production of nitric oxide (NO) and up to 10 times lower secretion of pro-inflammatory cytokines (IL-1𝛃 and TNF-α) in dendritic cells compared to macrophages. Moreover, DCs had a lower antimicrobial response to IFN-γ than bovine macrophages(51). NO was also measured in murine DCs exposed to M. bovis and M. bovis BCG; the results showed lower production of NO in the population infected with M. bovis compared to the one infected with BCG; however, NO production increased significantly when adding IFN-γ(52). Overall, these results suggest that DCs from these two species are permissive to M. bovis infection; however, IFN-γ only rescued NO production in murine DCs, evidencing a species-specific response.
Another study addressed the influence of bone marrow-derived DCs on the T lymphocyte profile in M. bovis infection in murine. Analyses of transcription levels, histopathology, and secretion molecules were carried out ten times during 56 d post-infection. The main findings were as follows: 1) Influence of high levels of prostaglandin-2 (PGE2) and cyclooxygenase-2 (COX2) mRNA on the cytokine profile (IL-17/IL-23); 2) Naïve LTCD4 were stimulated for differentiation towards Th17 and Treg, and 3) High bacterial load and tissue damage was observed in M. bovis infection. Considering these results, the researchers suggested that the induction of the PGE-2/COX-2 axis during infection with M. bovis contributes to sustained over-inflammation and could be related to the higher tissue damage(53). The greater permissiveness, higher response to external stimuli, and differentiation of T lymphocytes under M. bovis infection, could represent a key mechanism of very early immune modulation by the mycobacteria.
Lymphocyte function is important in M. bovis infection because TCD4+ lymphocytes produce IFN-γ that induces the microbicidal activity of macrophages and CD8+ T cells have also shown lytic activity on infected cells(54,55,56). A recent transcriptomic study compared whole blood from uninfected cattle and cattle experimentally infected with M. bovis at 8 and 20 wk post-infection. This study found that M. bovis infection upregulated chemokine genes such as monocyte chemoattractant protein 2 and chemokine (C-C motif) receptor 8 (CCR8), which are related to chemotaxis of monocytes and T lymphocytes, respectively, and downregulated genes related to class I antigen presentation and chemokines of neutrophils. The granzyme B gene was notably upregulated in the early and late stages of the infection, suggesting it may function as an infection biomarker. Since the genetic profile found high expression of cellular chemotactic genes and granzyme B, these are likely the most relevant defense mechanisms during the infection. In addition, the sustained transcription of granzyme B suggests that M. bovis antigens are being recognized by the population of cytotoxic T lymphocytes(57).
Although IFN-γ is a key cytokine in M. bovis infection, other circulating cytokines have been related to specific T lymphocyte populations. For example, T CD4+ lymphocytes and γδ T cells were identified as the main sources of IL-17 and IL-22, respectively, and a small population of γδ T cells produced both cytokines(58), besides a study in an experimental infection model found that the development of granulomas was directly related to increased IL-17 expression and decreased IL-22 expression. Therefore, the authors proposed IL-17 as a possible biomarker of bovine tuberculosis(59).
γδ T cells are particularly interesting, due to their production of IL-17 and also because this population is highly present in the circulation of bovines (up to 70 % in calves) compared to other species like humans and mice(60). The functions of γδ T cells in bovines include antigen presentation, IFN-γ production, cytotoxic activity, and regulation of the immune response has been reported(20,61). The genes expressed in subset WC1.1/T of γδ T cells from cows naturally infected with M. bovis were related to cell proliferation, activation, chemotaxis, and cytotoxic activity, evidencing their function on inflammation in bTB(62). A wider expression profile was described by quantifying mRNA from circulating γδ T cells and advanced-stage lung and lymph node granulomas. The analysis identified IFN-γ and IL-17 as the genes with the greatest differential expression between circulating γδ T cells of infected vs uninfected cattle. Furthermore, CCL2, IL-17, IL-10, and IFN-γ showed the greatest expression in the γδ T cells surrounding the granulomas(63). Overall, the production of chemoattractants, pro-inflammatory, and anti-inflammatory factors by circulating γδ T cells and those located in the infection site demonstrates their importance in the initial response and in maintaining the structure of the granulomatous lesion.
The series of cellular and molecular events induced by infection leads to the formation of granulomas, which are considered defense mechanisms against mycobacterial infections(64). In bTB, granulomas are classified into four stages(65) that have been used as a study reference(21,30). Previous work showed that granulomatous lesions in lung and mediastinal lymph nodes from naturally infected calves were devoid of capsules and displayed more necrosis and mycobacterial antigens than granulomas from adult cows(24). In addition, granulomatous tissue from calves showed more CD3+ positive cells and higher concentrations of TNF-α, IFN-γ, and inducible nitric oxide synthase (iNOS), as well as fewer γδ T cells compared to granulomas of adult cattle(66). These data suggest that age is a determining factor in the pathogenesis and immune response to bTB.
The humoral response to bTB was evaluated in 6-mo-old calves infected with different strains of M. bovis. The results identified antibodies against the antigens early secretory antigenic target (ESAT-6), culture filtrate protein (CFP10), and protein MPB83; however, the response was highly variable among animals and was predominant at week 18 post-infection. Moreover, antibodies directed against MPB83 remained constant from week 4 post-infection, regardless of the strain used(67). MPB83, MPB70, and ESAT-6/CFP10 were also evaluated in a comparative serological characterization performed in cattle, bison, and buffaloes naturally infected with M. bovis. In cattle, the predominant response was towards MPB70/MPB83; in bison, the response was similar towards the two antigenic groups; and in buffalo, the response was very low. Unlike ESAT-6/CFP10, which exclusively induces the production of IgG antibodies, MPB70/MPB83 were recognized by IgM and IgG antibodies. These results highlight the heterogeneity of the humoral response between species. Furthermore, the researchers hypothesized that M. bovis antigens induce the two antibody isotypes by reactivation at different times throughout the disease, which would explain the simultaneous presence of IgG and IgM(68).
Although most immunological studies in bovine tuberculosis have been directed to evaluate the response against infection using different strains of M. bovis, co-infection with other microorganisms has also been reported. For example with viruses(69), with other bacteria like, Brucella(70), and parasites(71-73). In most of the works where co-infection with M. bovis is reported, a statistical positive correlation with greater susceptibility and severity of bTB is suggested, however, studies with a functional approach at the cellular, molecular and tissue levels are necessary to elucidate the immunological dynamics and the effect on the evolution of bTB in the same host.
The diversity of immunological responses to M. bovis in vitro and in vivo models and the capacity of M. bovis to infect around 85 animal species(74) highlight its high capacity for adaptation and development of different immune evasion mechanisms. Considering all of the above, it suggests that these key variables strongly influence the outcome of the infection: 1) The age and breed of cattle; 2) The immune response to the infection, i.e., the greater permissiveness of some cells, the cell populations involved, the type of cell death, maturation stage of granulomatous lesions, and co-infection. Studying these variables through a comprehensive approach could generate more systematic knowledge to understand the high heterogeneity of bovine tuberculosis.
Cell death pathway in bovine tuberculosis
Apoptosis or programmed cell death
Apoptosis consists of a series of molecular processes known as programmed cell death(26). This concept was previously reported in silk moths(75), and the term apoptosis was only used until 1972(76). Research in this field has identified the genes involved in its initiation and regulation, which led to the award of the 2002 Nobel Prize in physiology(77). Currently, it is known that caspases (cysteine-aspartic acid proteases) are the initiating proteins of apoptosis in humans(78).
The morphological changes observed during apoptosis are cell shrinkage and a decrease in the nucleus size, characterized by Deoxyribonucleic acid (DNA) fragmentation, chromatin condensation, and detachment of cells from the surrounding tissue. Apoptotic bodies are also formed; these are phagocytosed by cells that arrive at the site due to the exposure of phosphatidylserine in the apoptotic cell membrane(79). Depending on the stimulus and the balance between an extensive group of pro- apoptotic and anti-apoptotic molecules, apoptosis can take two pathways: the intrinsic pathway (triggered by perturbations of the cell microenvironment, in particular, the mitochondria and endoplasmic reticulum) and the extrinsic pathway (induced by disturbances of the extracellular microenvironment and mediated by receptors)(79).
Some stimuli that activate the intrinsic mitochondrial pathway are hormones, radiation, toxins, hypoxia, and viral infections. These stimuli affect the permeability of the mitochondrial intermembrane(80), resulting in the release of pro-apoptotic proteins and cytochrome C to the cytoplasm. The interaction between apoptosis protease-activating factor-1 (Apaf-1) and caspase-9 forms the apoptosome, which activates the effector caspase 3. Furthermore, the Second Mitochondrial Activator of Caspases/Direct IAP-Binding Protein with Low pI (SMAC/DIABLO) inactivates an inhibitor of apoptosis factor (IAP). All molecular dynamics are regulated by proteins of the BCL-2 family of pro-apoptotic or anti-apoptotic nature, which are found in the cytoplasm and the outer membrane of the mitochondria(81,82).
Endoplasmic reticulum (ER) stress is associated with apoptosis. ER stress may be caused by loss of intracellular calcium balance, accumulation of misfolded proteins in the lumen of the ER, and disturbed protein transport to the Golgi apparatus(83). These conditions activate the unfolding protein response (UPR) system, composed of proteins such as inositol-requiring protein-1 (IRE1α) and protein kinase RNA (IPK-R)-like ER kinase (PERK), which activate accessory molecules or interact with each other to either restore balance or induce cell death(84). During a prolonged period of ER stress, the expression of pro-apoptotic proteins increases, and they interact with other molecules to promote apoptosis. For example, IRE1α activates apoptotic signaling-regulating kinase-1 (ASK1), which initiates a cascade of reactions that lead to the activation of pro-apoptotic molecules (Bim) and inactivation of anti-apoptotic molecules (Bcl-2)(85,86,87).
The extrinsic pathway of apoptosis is induced by receptor-ligand interactions. The most important ligands and receptors for apoptosis belong to the Tumor Necrosis Factor superfamily. Ligands can interact with one or more receptors, and most receptors are transmembrane proteins with an extracellular N-terminal that interacts with the ligand and an intracellular C-terminal that has a death domain(88). The interaction with this death domain can activate effector caspases through several pathways. For example, the FAS/FASL interaction along with adapter proteins can bind to pro-caspases 8 and 10 and subsequently activate effector caspases by autocatalysis(89) or form protein complexes that activate or inhibit caspases, as occurs with the TNF receptor(90,91).
Apoptosis is an essential mechanism to maintain cellular homeostasis(92-98) and also represents a defense mechanism in the immune response, especially against intracellular pathogens(99).
Role of apoptosis in Mycobacterium bovis infection
In mycobacterial infections, apoptosis has been associated with reduced bacterial spread and viability(1,2). However, virulent mycobacterial strains and antigens may inhibit apoptosis in cells infected previously(2,3).
Complete mycobacteria
One of the first publications reporting apoptosis in M. bovis-infected bovine macrophages showed that cell death occurred as early as 4 h post-infection using different multiplicities of infection (MOI). The authors concluded that apoptosis was time and MOI-dependent(5). Using the same cell model, apoptosis was enhanced by IFN-γ/LPS and diminished by blocking TNF-α. In addition, in the presence of IL-10, mycobacterial intracellular replication was inversely related to apoptosis, suggesting that apoptosis plays a protective role against infection(6).
The Rodriguez group compared mice previously infected with attenuated vs virulent M. bovis strains. They identified that the virulent strain had a greater capacity to inhibit apoptosis in alveolar macrophages. In addition the apoptosis was decreased by IL-10 and increased by TNF-α(7). The previous findings were carried out in vivo and in vitro in macrophages infected with M. bovis. This study demonstrates that mycobacteria modulate apoptosis through cytokine production, level of virulence, and exposure dose. In Table 1, there are some of the most relevant findings of apoptosis in infection with the main causal agent of bTB.
Natural resistance against a disease is defined as the ability of the host to resist the development of a disease after the first exposure to the pathogen and without prior immunization(8). Natural resistance to mycobacterial infections in cattle has been reported by several authors. For example, the Esquivel-Solis group(9), compared apoptosis and microbicidal activity in resistant and susceptible bovine macrophages infected with M. bovis. The findings indicate that apoptosis increased in macrophages with high NO levels, suggesting a relationship between apoptosis and microbicidal activity in the resistant phenotype(9). These results coincide with those obtained from resistant macrophages infected with M. tuberculosis(100,101). The effect of IL-4 was studied in bovine macrophages in both phenotypes. The results show a decrease in the expression of pro-inflammatory genes and a lower tendency towards apoptosis in resistant macrophages, evidencing that alternative activation by IL-4 increased susceptibility to infection in resistant macrophages(102). The relationship between NO production, apoptosis, and intracellular survival of mycobacteria was also evaluated in dendritic cells in mice. Apoptosis (DNA fragmentation and caspases 3, 6 and 9) and bacterial concentration were quantified in the absence or presence of an iNOS inhibitor. Results from this study showed that: a) the population infected with BCG showed more apoptosis compared to M. bovis, b) in the presence of the inhibitor, apoptosis was significantly reduced in both infected populations, and c) M. bovis survived better than BCG in DCs. These results suggest that the reduced production of NO by dendritic cells due to the infection with M. bovis modulates the development of apoptosis and increases the possibility of mycobacterial survival(52). These results highlight the role of nitric oxide in apoptosis in the early phases of infection.
Several research groups have focused on specific intracellular targets to understand the mechanisms and organelles involved in apoptosis. Vega, et al(103) in 2007 suggested an association between apoptosis and the nuclear translocation of Apoptosis-Inducing Factor (AIF) and mitochondrial membrane depolarization in macrophages exposed to an M. bovis protein extract(103). This prompted the investigation of other apoptosis-associated components, for example, the impact of mitochondrial permeability on DNA fragmentation and mycobacterial viability in bovine macrophages infected with M.bovis. DNA fragmentation decreased independently of caspase activity when mitochondrial permeability was inhibited. Furthermore, the translocation of AIF and Endonuclease G (Endo-G) to the nucleus, measured by immunoblot, increased 15 and 43 times, respectively, and the viability of the intracellular mycobacteria increased by 26 %(104). These results support the idea that the translocation of Endo-G to the nucleus is also involved in DNA fragmentation as a result of M. bovis infection by altering mitochondrial permeability. The identification of molecules such as Endo G and AIF in the nucleus and the decreased intracellular mycobacterial viability in the absence of activated caspases, suggest that caspase activation is not required for DNA fragmentation and reveals the existence of different mechanisms in the development and modulation of apoptosis, especially during infection.
Mitochondrial stress induced by M. bovis infection was also evaluated in THP-1 cells. It was found that apoptotic caspases negatively modulate IFN-β production by reducing the nuclear translocation of p-IRF3 (Interferon Regulatory Factor 3)(105). This represents a beneficial scenario for the host since a lower IFN-β in M. bovis infection has been associated with a better prognosis(106).
The endoplasmic reticulum stress induced by mycobacterial infection was investigated in murine macrophages previously infected with M. bovis. This study showed a higher intracellular survival of mycobacteria upon adding an ER stress inhibitor, which directly modulated the percentage of apoptotic cells(107). The relationship between apoptosis and the functionality of the mitochondria and ER and its impact on intracellular mycobacterial viability highlights the protective effect of apoptosis against infection(108). However, these organelles and pathways can also become targets for mycobacteria modulation.
Activated caspases have been used as the only marker of apoptosis in mycobacterial infections(109). Nevertheless, caspase-independent apoptosis has been identified in cattle and buffaloes infected with M. bovis(103,110). Furthermore, since apoptosis limits the intracellular growth of mycobacteria, the absence of caspase activation could represent a mechanism of evasion of cell death by M. bovis(107).
Most approaches to investigating apoptosis in mycobacterial infection have been carried out in cell models (mainly macrophages), allowing the study of the protective role of apoptosis in the acute phase. However, due to the persistent nature of the infection, apoptosis should also be studied in the chronic phase of mycobacterial infection. The Cherdantseva group reported that apoptotic cells corresponded to approximately 11 % of cells in lung granulomas of mice infected with M. bovis-BCG after 180 days of infection. These data suggest that, although apoptosis is induced at the tissue level, it is insufficient to eliminate the mycobacteria during the development of the pathological process(111).
Proteins from M.bovis
The dual role of apoptosis in mycobacterial infection has been observed in studies using individual M. tuberculosis antigens, in which the antigens are classified as pro-apoptotic or anti-apoptotic. These studies suggest that mycobacteria modulate the cell death mechanism through dynamics of antigenic expression(2). In this context, to identify specific M. bovis apoptosis-inducing proteins, bovine macrophages were exposed to different protein extract fractions. Caspase-independent apoptosis was induced by two M. tuberculosis recombinant proteins, hsp70 and heparin-binding haemagglutinin (HBHA), which have high homology with M. bovis in bovine macrophages(112). In this regard, efforts have been made to determine the protein profile in the protein extracts of M. bovis. Using mass spectrophotometry, MPB70, MPB83, and 60-kDa chaperonin were identified as the main protein candidates that induce caspase-independent apoptosis(28).
M. bovis and M tuberculosis have a high genomic homology(113). Therefore, investigating the modulatory effect exerted by highly homologous proteins in the two species could identify the new mechanisms in cell death. This knowledge would allow to understand the particularities of the infection and the general pathogenesis of bTB.
All together, the above mentioned results indicate that apoptosis is a multifactorial event involving characteristics of the bacteria (such as virulence, time and multiplicity of infection) and intrinsic characteristics of the affected host. However, despite the effect of these multiple variables, apoptosis and intracellular growth of mycobacteria in bovine macrophages are inversely correlated, which suggests that apoptosis in M. bovis infection represents a host defense mechanism.
Necrosis or accidental cell death
The term necrosis comes from the Greek "necro," which means corpse or death and “osis”, which means condition or state. Necrosis was used to describe the morphological death of cells resulting from infection, cell damage, noxious stimuli, or mechanical damage; therefore, necrosis was thought to be due to abrupt changes leading to accidental cell death(114). Pathological diagnosis evaluates macroscopic and microscopic features in the affected tissue and classifies necrosis as coagulative, fibrinoid, hemorrhagic, and caseating(115).
In M. bovis infection, necrosis is present in the advanced stages of granulomatous lesions(65). In addition, unregulated necrosis has been associated with a higher spread of mycobacterial infection(116). An analysis of granulomas from cattle naturally infected with M. bovis showed large necrotic areas with central calcification, no connective tissue capsule, and few giant cells. Necrosis was the predominant cell death observed, and it was accompanied by more mycobacterial antigens, which was mainly observed in calves(24). Moreover, inducing necrosis using hydrogen peroxide in M. tuberculosis-infected macrophages favors the escape of the bacteria to the extracellular medium without affecting its viability(117). In addition, other necrosis induction assays allow the exit and proliferation of mycobacteria(118) .
Different types of regulated cell death
In recent decades, biochemical and molecular advances have enabled the discovery of some types of necrosis that are not accidental, rather, they follow regulated signaling pathways that produce a necrotic morphology(99,100,119,120). The description of these signaling pathways has helped define the diverse pathways of cell demise that lead to necrotic cell death. Among the different types of necrotic cell death are necroptosis, pyroptosis, among others(99,101,102,103,119,121,122,123).
Pyroptosis
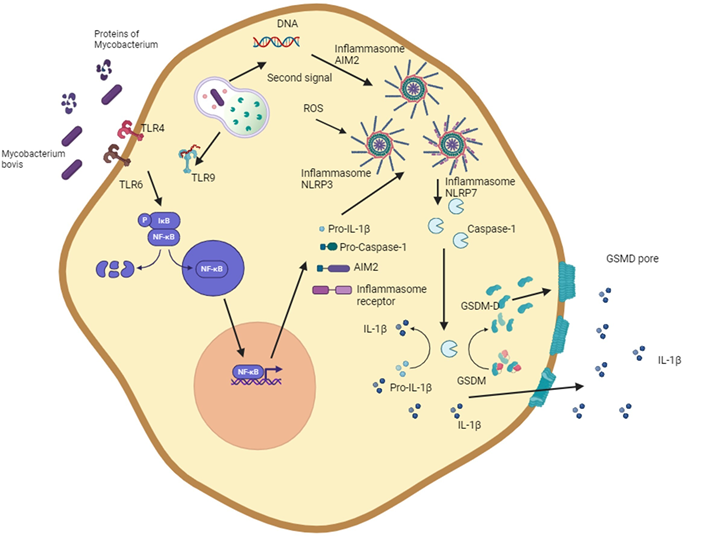
The term pyroptosis is derived from the Greek “pyro” (fire, fever) and “ptosis” (falling)(104). Pyroptosis was first described in Salmonella and Shigella in vitro infection models, in which caspase 1 initiated cell death(104,105,106,124,125,126). Pyroptosis is an inflammatory cell death classically characterized by the inflammasome, caspase 1, gasdermin D (GSDMD), and the release of IL-1β and IL18 (Figure 1).
Figure 1: Routes of induction of pyroptosis by Mycobacterium bovis
The diagram shows the ability of M. bovis and mycobacterial proteins to activate NLRP3 and AIM2 inflammasomes. The activation of the NLRP3 inflammasome is initiated through pattern-recognition receptors and then by multiple stimuli such as the generation of reactive oxygen species, potassium efflux, or lysosomal components. Activation of the AIM2 inflammasome is initiated by bacterial DNA recognition. The assembly of the inflammasome leads to the maturation of IL-1β and the cleavage of gasdermin, forming gasdermin D which damages the cell membrane and results in necrotic cell death. This figure was created using BioRender.com.
The inflammasome, which becomes activated in pyroptosis, consists of multiprotein structures including a receptor of the NLR (nucleotide-binding oligomerization domain-like receptors) or AIM myeloma 2 (AIM2)-like receptors families, as well as the ASC (Apoptosis-associated speck-like protein containing a CARD) and pro-caspase 1(107,108,127,128). However, less frequently, pyroptosis can be activated by an alternative pathway. Activation of the inflammasome leads to the activation of inflammatory caspases (caspase-1,-4,-5 in humans and caspase-1 and -11 in mice) and the cleavage of the interleukin-1 family and GSDMD. The active GSDMD can assemble to form pores in the cell membrane and generate an osmotic imbalance that leads to cell death under an inflammatory environment(109,110, 129,130).
M. bovis can induce pyroptosis in macrophage cells and macrophage-derived cell lines (Table 1). The strain of the bacterium, the multiplicity of infection, and the time after infection are among the factors that favor pyroptosis(10,131,132,133). The main mechanisms that induce pyroptosis are related to the canonical activation of inflammasomes (Figure 1). NLRP7, which recognizes bacterial glycoproteins; AIM2, which recognizes double stranded DNA; and NLRP3, which is activated by various signals, such as potassium efflux, ROS, extracellular ATP, pore-forming toxins, and mediate pyroptosis associated to M. bovis infection(111,112,113,131,132,133). The activation of inflammasomes affects the production of IL-1β, IL-18, and IL-33, generating an inflammatory environment that helps control the infection produced by mycobacteria(132).
Inflammasome NLRP3 activation requires two signals and generates an inflammatory environment. Stimulation of macrophages with LPS increases IL-1 β and nitric oxide, which may limit the intracellular growth of mycobacteria(9,10). Activating the inflammasome by M. bovis-infected bovine macrophages decreases the intracellular growth of mycobacteria(10).
The inflammatory environment generated by pyroptosis can regulate the proliferation of bacteria, recruiting immune cells that help control bacterial infections. However, pyroptosis can cause tissue damage, therefore, it represents a strong mechanism that some host cells have to control bacterial intracellular growth. Of note, there is currently no information on which bacteria strains commonly induce pyroptosis. It is also unknown whether bacterial growth is controlled or whether some bacteria induce this type of cell death to escape from the cells and infect the surrounding tissues(12,134).
Autophagy
The term autophagy is derived from the Greek “auto” (self) and “phagen” (to eat). Autophagy is a highly conserved pathway that degrades cellular components using lysosomes(135). Autophagy is a regulated mechanism that allows cells to survive under nutrient deprivation or adverse conditions. However, autophagy can also cause cell death (autophagy-cell death dependent). This mechanism can occur concomitantly with another type of cell death, such as apoptosis , or start as autophagy and trigger apoptosis(136).
Autophagy has been shown to limit intracellular bacteria. Some of the molecules involved in this process are myeloid-related protein 8/14 and interferon-γ inducible protein 204 (IFI204) that induces autophagy in peripheral blood mononuclear cells and THP1 cells in a ROS-dependent manner, which inhibits the intracellular growth of Mycobacterium BCG (Table 1)(137). Moreover, IFI204 is a DNA sensor that activates the innate immune response, including autophagy and interferon-β production (IFN-β). IFI204 proteins are involved in IFN-β responses by recruiting STING to activate TBK-1-IRF3 pathways. Induction of autophagy by IFI204 induces phosphorylation of TBK-1 to inhibit M. bovis survival in macrophages(138).
Importantly, M.bovis can evade autophagy. One of the mechanisms consists in the specific inhibition of autophagy responsible for the control of intracellular organisms (xenophagy), for example, through the activation of the PINK1-PRKN/Parkin indicating pathway involved in mitophagy, which generates a competition of both pathways for p-TBK1 leading to a decrease in xenophagy and the survival of the mycobacteria(139). The role of the microRNA miR-199a was evaluated in macrophages derived from bone marrow, lung, and spleen of M. bovis-infected mice. The infection increased the expression of miR-199a, and this suppressed autophagy by blocking phagolysosome maturation through the interaction with TANK binding Kinase 1. These changes led to an increase in intracellular survival of the mycobacteria. These results provide a mechanism for M. bovis to evade elimination(137).
Although the development of autophagy participates in maintaining cellular balance, it may also function as an innate immune response mechanism that limits the growth of intracellular bacteria. In infections with M. bovis, autophagy is induced by low-virulence bacteria, suggesting that M. bovis may also modify processes involved in sustaining cellular homeostasis(140,141).
Conclusions
Regardless of the influence of different variables (such as virulence, time, species, and the host resistance phenotype) on apoptosis, experimental results suggests that cell death by apoptosis helps to control bacterial growth.
The bacterial inhibitory effect on apoptosis, the redirection of autophagy, and the induction of inflammatory cell death such as necrosis and pyroptosis may be bacterial mechanisms to evade the host immune response.
Although experimental conditions allow the detection of a specific type of cell death, the simultaneous activation of multiple types of cell death, known as PANoptosis, has also been observed in M. tuberculosis infection. This scenario opens the possibility of studying M. bovis infection in a global manner that considers all experimental variables and phases of the different cell death types.
The high adaptability of M. bovis and the key role of cell death in immune activation highlight the need for more studies on regulated and non-regulated cell death. These studies will increase the understanding of bovine infection and aid in developing new strategies to counteract bovine tuberculosis.
The most important points of this review can be numbered in: 1) Cell death by apoptosis helps to control bacterial growth. 2) Autophagy is a conserved mechanism that limits mycobacterium intracellular replication. 3) Pyroptosis is an extreme mechanism that helps control M. bovis at the cost of damaging host tissue. 4) Necrosis will allow the escape and proliferation of mycobacteria.
Acknowledgements
We thank the DGAPA UNAM Postdoctoral fellowship for Paola Andrea Ortega Portilla as recipient. Omar Escobar-Chavarría received support from a Consejo Nacional de Humanidades, Ciencia y Tecnología CONAHCYT scholarship. This research was funded by a grant from the Universidad Nacional Autónoma de México, DGAPA-PAPIIT IG201521 and DGAPA-PAPIIT IG200918.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Table 1: Cell death in Mycobacterium bovis infection
+Target molecule: molecule selected to evaluate cell death, (R): Resistance phenotype, ND: Not Determined, Mycobacterial load*: Quantified in the presence of the specific type of death concomitantly, AIF: Apoptosis Inductor Factor, HBHA: heparin-binding haemaglutinin, AIM2: absent in melanoma 2, LDH: lactate dehydrogenase, NLRP3: NOD, leucine-rich repeats and pyrin domain-containing protein 3, NOD, leucine-rich repeats and pyrin domain-containing protein 7, LC3: Microtubule-associated protein 1A/1B-light chain 3, LAMP-1: lysosomal associated membrane protein 1, HSPD1: heat shock 60-kDa protein 1, PPE: Pro-Glu motif-containing (PE) and Pro-Pro-Glu motif-containing (PPE) family proteins, BMDM: bone-marrow derived macrophages, MRP8/14: Myeloid-related proteins (MRPs) 8 and 14, LRG-47: IFN-inducible protein Irgm1.
Literature cited: