https://doi.org/10.22319/rmcp.v16i1.6654
Revisión bibliográfica
Control y diseminación bacteriana asociada a la muerte celular en la infección por Mycobacterium bovis. Revisión
Paola Andrea Ortega-Portilla a
Omar Escobar-Chavarría b
José Ángel Gutiérrez-Pabello a*
a Universidad Nacional Autónoma de México. Facultad de Medicina Veterinaria y Zootecnia. Departamento de Microbiologia e Inmunologia. Laboratorio de Investigación en Tuberculosis y Brucelosis. Ciudad de México, México.
b Universidad Nacional Autónoma de México. Facultad de Estudios Superiores Cuautitlán. Estado de México, México.
*Autor de correspondencia: jagp@unam.mx
Resumen:
Una de las características distintivas de la infección por Mycobacterium bovis es la muerte celular. El tipo de muerte celular que se produce durante la infección determina la persistencia de las enfermedades micobacterianas. El objetivo de este artículo es proporcionar una revisión integral y trazar los posibles escenarios de los tipos de muerte celular en la patogénesis de la tuberculosis bovina. Con los datos actuales se puede concluir que: 1) el desarrollo de la apoptosis y sus diferentes variantes está relacionado con el control de micobacterias, 2) la autofagia es un mecanismo conservado que limita la replicación intracelular de micobacterias, 3) la piroptosis es un mecanismo extremo que ayuda a controlar M. bovis a costa de dañar el tejido del hospedero, y 4) la necrosis permitirá el escape y proliferación de micobacterias.
Palabras clave: Muerte celular, Tuberculosis bovina, Mycobacterium bovis, Apoptosis, Autofagia, Piroptosis.
Recibido: 06/03/2024
Aceptado: 30/09/2024
Introducción
La forma en que muere una célula juega un papel crucial en los procesos fisiológicos. En las infecciones por micobacterias, algunos tipos de muerte celular han sido catalogados como mecanismos de defensa del hospedero, pero también como consecuencia de los factores de virulencia del patógeno(1,2,3). El Comité de Nomenclatura de Muerte Celular (NCCD, por sus siglas en inglés) 2018, propone clasificar los tipos de muerte celular con base en los aspectos mecanicistas y esenciales del proceso, categorizando la mayoría dentro del grupo de muerte celular regulada(4). De este gran grupo, algunos tipos han sido reportados en infecciones por micobacterias, por ejemplo, apoptosis(5-9), piroptosis(10), ferroptosis(11) y necroptosis(12).
El Mycobacterium bovis (M. bovis) pertenece al complejo Mycobacterium tuberculosis. Esta especie es el agente causal de la tuberculosis zoonótica y el principal agente etiológico de la tuberculosis bovina (TBb). M. bovis afecta a muchas especies animales(13,14), por lo que es un problema para la salud pública y el sector ganadero(15,16).
M. bovis se transmite principalmente por aire, a través de las gotitas exhaladas del sistema respiratorio de los animales infectados. Se desarrolla una respuesta inmune celular, que es considerada el principal mecanismo inmune contra las bacterias intracelulares(17,18). La dinámica entre macrófagos, neutrófilos, fibroblastos, células dendríticas, linfocitos B, linfocitos T γδ, linfocitos CD4+, CD8+ y citocinas proinflamatorias como el factor de necrosis tumoral (TNF) y el interferón gamma (IFN-γ), dan lugar a la formación de la estructura de defensa característica contra las micobacterias: el granuloma(19,20,21).
Las lesiones granulomatosas son características de la TBb, y se encuentran principalmente en los ganglios linfáticos y en los pulmones(22). Su desarrollo varía en los diferentes ganglios linfáticos de un mismo animal(23) y, además, se ha reportado variación en la morfología estructural de granulomas de terneros y adultos(24), por lo que se considera que los granulomas en la TBb tienen una presentación heterogénea.
La muerte celular es uno de los mecanismos determinantes en la formación y evolución del granuloma que impulsa el desarrollo de la infección y la presentación de la enfermedad(25). Como consecuencia de la persistencia de la infección por M. bovis, pueden ocurrir varios tipos de muerte celular, destacándose la apoptosis y la necrosis(5-9,25). Sin embargo, otras modalidades, como la piroptosis y la autofagia, también pueden desempeñar un papel en la infección(10,26,27,28). Esta revisión tiene como objetivo proporcionar un resumen integral de los tipos de muerte celular que se han identificado en la infección por M. bovis y resaltar su impacto en el hospedero. Para lograr este objetivo, se presenta la información dividida en dos secciones principales: 1) Patogénesis y respuesta inmune en la TBb y 2) Vías de muerte celular en la TBb.
Patogénesis y respuesta inmune en la tuberculosis bovina
La tuberculosis bovina se transmite por contacto directo con productos de excreción infectados (orina, saliva, leche, semen, secreciones uterinas) o micobacterias presentes en gotitas exhaladas por el sistema respiratorio de animales infectados(29). El sistema respiratorio se ve afectado principalmente, incluyendo los pulmones y los ganglios linfáticos asociados(24,30,31). Las lesiones en el aparato digestivo se han relacionado con la transmisión por ingestión de alimentos contaminados(32), y la transmisión transplacentaria ocurre en terneros nacidos con infección congénita(33).
La TBb puede ser subclínica por largos períodos, sintomática (fiebre, pérdida de peso, dificultad respiratoria y disminución de la producción de leche), o tener una evolución hacia una presentación generalizada como consecuencia de la diseminación linfática o hematógena de las micobacterias, cambiando a las fases temprana y tardía de la infección(14). Factores como la localización de la enfermedad, la evolución de la lesión primaria, el factor de virulencia micobacteriana, la concentración bacteriana, el desarrollo de lesiones granulomatosas y la inmunocompetencia del hospedero, son determinantes para la presentación de síntomas clínicos(15,21,29,34,35).
La respuesta inmune juega un papel crucial en la evolución de la infección en las fases aguda y crónica(36). En particular, las respuestas mediadas por células son vitales(37). Dado que el sistema respiratorio es uno de los más afectados por M. bovis, se han realizado estudios transcripcionales y funcionales en diferentes poblaciones celulares de este sistema.
Los macrófagos alveolares se encuentran entre las primeras poblaciones celulares infectadas por micobacterias inhaladas, por lo que se han estudiado utilizando diferentes enfoques. Los análisis transcriptómicos han revelado que los cambios en la expresión génica son contrastantes. Por ejemplo, se ha reportado una disminución en la expresión de genes relevantes para el reconocimiento de M. bovis(37,38) y una mayor polarización de los macrófagos hacia un fenotipo M2 más permisivo-replicativo(39). Por otro lado, los genes que codifican quimiocinas, receptores de reconocimiento y moléculas proinflamatorias mostraron un aumento tras la infección por M. bovis(40,41). Y, finalmente, este enfoque también identificó variación genómica relacionada tanto con la susceptibilidad como con la resistencia a la infección(42). Otro estudio que utilizó análisis de composición y metabolismo lipídico identificó diferencias significativas en el grupo lipídico entre M. tuberculosis relacionadas con la formación de macrófagos espumosos y M. bovis con inhibición de la autofagia(43).
Estos hallazgos relacionados con la función protectora y no protectora de los macrófagos alveolares frente a la infección, acompañada de una respuesta dependiente de las especies micobacterianas, demuestran el papel determinante que juega la interacción micobacteria/macrófago alveolar tanto en la fase aguda como en la evolución de la infección.
Los neutrófilos son otra población celular importante en la infección por micobacterias(44). Los neutrófilos bovinos funcionan como células reguladoras principalmente en la inmunidad innata de bovinos clínicamente sanos, pero también en condiciones de infección(45). Por ejemplo, los neutrófilos bovinos expuestos a M. bovis aumentaron la fagocitosis, la activación celular, la secreción de citocinas proinflamatorias y la replicación intracelular(46). Estos resultados sugieren que la infección por M. bovis podría modular la respuesta en neutrófilos bovinos.
La β-defensina-5 es un péptido antimicrobiano almacenado en macrófagos bovinos y gránulos de neutrófilos. La incubación de β-defensina-5 recombinante con M. bovis evidenció sus efectos antimicrobianos dependientes del tiempo; este péptido inhibió el crecimiento en un 88 % y alteró la pared micobacteriana a las 72 h de incubación(47). También se demostró el papel inmunoprotector de la β-defensina-5 recombinante. La β-defensina-5 recombinante de neutrófilos bovinos se utilizó en la inmunización de ratones, que luego fueron infectados con M. bovis. Los resultados mostraron una reducción del tejido inflamatorio y de la carga bacteriana en el pulmón y el bazo, demostrando el potencial de su función inmunoprotectora(48).
Los cambios en la estructura de los núcleos de neutrófilos han sido sugeridos como un método diagnóstico complementario para la tuberculosis bovina. En neutrófilos humanos expuestos a suero de vacas con Derivado Proteico Purificado de Mycobacterium (PPD+), después de 3 h, la pinocitosis fue el cambio más común observado en los núcleos celulares(49). Además, el patrón específico de expresión de los genes transcripcionales inducibles por IFN, los genes de la mieloperoxidasa (MPO) y los genes de la proteína relacionada con la pentraxina (PTX3), de neutrófilos, mostraron su potencial como herramientas de diagnóstico para la infección por M. bovis en el ganado(50). A pesar del efecto modulador que aparentemente ejerce M. bovis sobre los neutrófilos, los hallazgos antimicrobianos de algunas de sus moléculas intracelulares evaluadas de manera recombinante podrían representar un campo de investigación para el desarrollo biotecnológico con potencial para su aplicación en diagnóstico y terapéutica.
Teniendo en cuenta la importancia de las células dendríticas (CD) en la inmunidad innata y adaptativa, algunos grupos de investigación han estudiado su respuesta frente a la infección por M. bovis. Un análisis comparativo entre CD y macrófagos bovinos, ambos infectados con M. bovis, identificó una menor producción de óxido nítrico (NO) y una secreción hasta 10 veces menor de citocinas proinflamatorias (IL-1β y TNF-α) en células dendríticas en comparación con los macrófagos. Además, las CD tuvieron una menor respuesta antimicrobiana al IFN-γ que los macrófagos bovinos(51). También se midió el NO en CD murinas expuestas a M. bovis y M. bovis BCG; los resultados mostraron una menor producción de NO en la población infectada con M. bovis en comparación con la infectada con BCG; sin embargo, la producción de NO aumentó significativamente al agregar IFN-γ(52). En general, estos resultados sugieren que las CD de estas dos especies son permisivas a la infección por M. bovis; no obstante, el IFN-γ solo rescató la producción de NO en las CD murinas, lo que evidencia una respuesta específica de la especie.
Otro estudio abordó la influencia de las CD derivadas de la médula ósea en el perfil de linfocitos T en la infección por M. bovis en murinos. Los análisis de los niveles de transcripción, histopatología y moléculas de secreción se llevaron a cabo diez veces durante 56 días después de la infección. Los principales hallazgos fueron los siguientes: 1) influencia de altos niveles de ARNm de prostaglandina-2 (PGE2) y ciclooxigenasa-2 (COX2) en el perfil de citocinas (IL-17/IL-23); 2) se estimularon los LTCD4 naive para diferenciarlos hacia Th17 y Treg, y 3) se observó una alta carga bacteriana y daño tisular en la infección por M. bovis. Teniendo en cuenta estos resultados, los investigadores sugirieron que la inducción del eje PGE-2/COX-2 durante la infección por M. bovis contribuye a la sobreinflamación sostenida y podría estar relacionada con el mayor daño tisular(53). La mayor permisividad, mayor respuesta a estímulos externos y diferenciación de linfocitos T bajo infección por M. bovis, podrían representar un mecanismo clave de modulación inmune muy temprana por parte de las micobacterias.
La función de los linfocitos es importante en la infección por M. bovis porque los linfocitos TCD4+ producen IFN-γ que induce la actividad microbicida de los macrófagos y los linfocitos T CD8+ también han mostrado actividad lítica en las células infectadas(54,55,56). Un estudio transcriptómico reciente comparó sangre entera de ganado no infectado y ganado infectado experimentalmente con M. bovis a las 8 y 20 semanas después de la infección. Este estudio encontró que la infección por M. bovis reguló al alza los genes de quimiocina, como la proteína quimioatrayente 2 de monocitos y el receptor 8 (CCR8) de quimiocinas (motivo C-C), que están relacionados con la quimiotaxis de monocitos y linfocitos T, respectivamente, y reguló a la baja los genes relacionados con la presentación de antígenos de clase I y las quimiocinas de los neutrófilos. El gen de la granzima B se reguló notablemente al alza en las etapas tempranas y tardías de la infección, lo que sugiere que puede funcionar como un biomarcador de infección. Dado que el perfil genético encontró una alta expresión de genes quimiotácticos celulares y granzima B, estos son probablemente los mecanismos de defensa más relevantes durante la infección. Además, la transcripción sostenida de la granzima B sugiere que los antígenos de M. bovis están siendo reconocidos por la población de linfocitos T citotóxicos(57).
Aunque el IFN-γ es una citocina clave en la infección por M. bovis, otras citocinas circulantes se han relacionado con poblaciones específicas de linfocitos T. Por ejemplo, los linfocitos T CD4+ y los linfocitos T γδ fueron identificados como las principales fuentes de IL-17 e IL-22, respectivamente, y una pequeña población de linfocitos T γδ produjo ambas citocinas(58); además, un estudio en un modelo experimental de infección encontró que el desarrollo de granulomas estaba directamente relacionado con el aumento de la expresión de IL-17 y la disminución de la expresión de IL-22. Por lo tanto, los autores propusieron la IL-17 como un posible biomarcador de tuberculosis bovina(59).
Los linfocitos T γδ son particularmente interesantes, debido a su producción de IL-17 y también porque esta población está muy presente en la circulación de los bovinos (hasta el 70 % en los terneros) en comparación con otras especies, como los humanos y los ratones(60). Se han reportado las funciones de los linfocitos T γδ en bovinos, como la presentación de antígenos, la producción de IFN-γ, la actividad citotóxica y la regulación de la respuesta inmune(20,61). Los genes expresados en el subconjunto WC1.1/T de linfocitos T γδ de vacas infectadas naturalmente con M. bovis se relacionaron con la proliferación celular, activación, quimiotaxis y actividad citotóxica, evidenciando su función sobre la inflamación en TBb(62). Se describió un perfil de expresión más amplio mediante la cuantificación del ARNm de los linfocitos T γδ circulantes y los granulomas de pulmón y ganglios linfáticos en etapa avanzada. El análisis identificó el IFN-γ y la IL-17 como los genes con la mayor expresión diferencial entre los linfocitos T γδ circulantes del ganado infectado frente al no infectado. Además, CCL2, IL-17, IL-10 e IFN-γ mostraron la mayor expresión en los linfocitos T γδ que rodean los granulomas(63). En general, la producción de factores quimio-atrayentes, proinflamatorios y antiinflamatorios por parte de los linfocitos T γδ circulantes y los localizados en el sitio de la infección demuestra su importancia en la respuesta inicial y en el mantenimiento de la estructura de la lesión granulomatosa.
La serie de eventos celulares y moleculares inducidos por la infección conduce a la formación de granulomas, que son considerados mecanismos de defensa contra las infecciones por micobacterias(64). En la TBb, los granulomas se clasifican en cuatro estadios(65) que han sido utilizados como referencia del estudio(21,30). Un trabajo previo mostró que las lesiones granulomatosas en los ganglios linfáticos pulmonares y mediastínicos de terneros naturalmente infectados estaban desprovistas de cápsulas y mostraban más necrosis y antígenos micobacterianos que los granulomas de vacas adultas(24). Además, el tejido granulomatoso de terneros mostró más células CD3+ positivas y mayores concentraciones de TNF-α, IFN-γ y óxido nítrico sintasa inducible (iNOS), así como menos células T γδ en comparación con los granulomas de bovinos adultos(66). Estos datos sugieren que la edad es un factor determinante en la patogénesis y la respuesta inmunitaria a la TBb.
Se evaluó la respuesta humoral a la TBb en terneros de 6 meses de edad infectados con diferentes cepas de M. bovis. Los resultados identificaron anticuerpos contra los antígenos diana de secreción temprana (ESAT-6), la proteína de filtrado de cultivo (CFP10) y la proteína MPB83; sin embargo, la respuesta fue muy variable entre los animales y fue predominante en la semana 18 después de la infección. Además, los anticuerpos dirigidos contra MPB83 se mantuvieron constantes a partir de la semana 4 después de la infección, independientemente de la cepa utilizada(67). MPB83, MPB70 y ESAT-6/CFP10 también se evaluaron en una caracterización serológica comparativa realizada en bovinos, bisontes y búfalos infectados naturalmente con M. bovis. En bovinos, la respuesta predominante fue hacia MPB70/MPB83; en bisontes, la respuesta fue similar hacia los dos grupos antigénicos; y en búfalos, la respuesta fue muy baja. A diferencia de ESAT-6/CFP10, que induce exclusivamente la producción de anticuerpos IgG, MPB70/MPB83 fueron reconocidos por anticuerpos IgM e IgG. Estos resultados ponen de manifiesto la heterogeneidad de la respuesta humoral entre especies. Además, los investigadores plantearon la hipótesis de que los antígenos de M. bovis inducen los dos isotipos de anticuerpos por reactivación en diferentes momentos a lo largo de la enfermedad, lo que explicaría la presencia simultánea de IgG e IgM(68).
Aunque la mayoría de los estudios inmunológicos en tuberculosis bovina se han dirigido a evaluar la respuesta frente a la infección utilizando diferentes cepas de M. bovis, también se ha reportado la coinfección con otros microorganismos. Por ejemplo, con virus(69), con otras bacterias como Brucella(70) y parásitos(71-73). En la mayoría de los trabajos donde se reporta coinfección con M. bovis, se sugiere una correlación estadísticamente positiva con mayor susceptibilidad y severidad de la TBb, no obstante, son necesarios estudios con abordaje funcional a nivel celular, molecular y tisular para dilucidar la dinámica inmunológica y el efecto sobre la evolución de la TBb en el mismo hospedero.
La diversidad de respuestas inmunológicas a modelos in vitro e in vivo de M. bovis y la capacidad de M. bovis para infectar alrededor de 85 especies animales(74) ponen de manifiesto su alta capacidad de adaptación y desarrollo de diferentes mecanismos de evasión inmune. Teniendo en cuenta todo lo anterior, sugiere que estas variables clave influyen fuertemente en el resultado de la infección: 1) la edad y raza del ganado; 2) la respuesta inmune a la infección, es decir, la mayor permisividad de algunas células, las poblaciones celulares involucradas, el tipo de muerte celular, el estado de maduración de las lesiones granulomatosas y la coinfección. El estudio de estas variables a través de un enfoque integral podría generar un conocimiento más sistemático para comprender la alta heterogeneidad de la tuberculosis bovina.
Vía de muerte celular en la tuberculosis bovina
Apoptosis o muerte celular programada
La apoptosis consiste en una serie de procesos moleculares conocidos como muerte celular programada(26). Este concepto fue reportado previamente en polillas de seda(75), y el término apoptosis solo fue utilizado hasta 1972(76). Las investigaciones en este campo han identificado los genes implicados en su iniciación y regulación, lo que ha llevado a la concesión del Premio Nobel de Fisiología 2002(77). Actualmente, se sabe que las caspasas (proteasas de cisteína-ácido aspártico) son las proteínas iniciadoras de la apoptosis en humanos(78).
Los cambios morfológicos observados durante la apoptosis son la contracción celular y la disminución del tamaño del núcleo, caracterizados por la fragmentación del ácido desoxirribonucleico (ADN), la condensación de la cromatina y el desprendimiento de las células del tejido circundante. También se forman cuerpos apoptóticos; estos son fagocitados por las células que llegan al sitio debido a la exposición de la fosfatidilserina en la membrana celular apoptótica(79). Dependiendo del estímulo y del equilibrio entre un grupo extenso de moléculas proapoptóticas y antiapoptóticas, la apoptosis puede tomar dos vías: la vía intrínseca (desencadenada por perturbaciones del microambiente celular, en particular, las mitocondrias y el retículo endoplásmico) y la vía extrínseca (inducida por alteraciones del microambiente extracelular y mediada por receptores)(79).
Algunos estímulos que activan la vía mitocondrial intrínseca son las hormonas, la radiación, las toxinas, la hipoxia y las infecciones virales. Estos estímulos afectan la permeabilidad de la intermembrana mitocondrial(80), resultando en la liberación de proteínas proapoptóticas y citocromo C al citoplasma. La interacción entre el factor activador de la proteasa-1 de la apoptosis (Apaf-1) y la caspasa-9 forma el apoptosoma, que activa la caspasa efectora 3. Además, el Segundo Activador Mitocondrial de Caspasas/Proteína de Unión Directa a IAP con pI Bajo (SMAC/DIABLO) inactiva un inhibidor del factor de apoptosis (IAP). Toda la dinámica molecular está regulada por proteínas de la familia BCL-2 de naturaleza proapoptótica o antiapoptótica, que se encuentran en el citoplasma y en la membrana externa de las mitocondrias(81,82).
El estrés del retículo endoplásmico (RE) se asocia con la apoptosis. El estrés del RE puede ser causado por la pérdida del equilibrio de calcio intracelular, la acumulación de proteínas mal plegadas en el lumen del RE y la alteración del transporte de proteínas al aparato de Golgi(83). Estas condiciones activan el sistema de respuesta a proteínas desplegadas (UPR), compuesto por proteínas como la proteína-1 que requiere inositol (IRE1α) y la proteína quinasa ARN (IPK-R) similar al RE quinasa (PERK), que activan moléculas accesorias o interactúan entre sí para restaurar el equilibrio o inducir la muerte celular(84). Durante un período prolongado de estrés del RE, la expresión de proteínas proapoptóticas aumenta e interactúan con otras moléculas para promover la apoptosis. Por ejemplo, IRE1α activa la quinasa reguladora de señalización apoptótica 1 (ASK1), que inicia una cascada de reacciones que conducen a la activación de moléculas proapoptóticas (Bim) y la inactivación de moléculas antiapoptóticas (Bcl-2)(85,86,87).
La vía extrínseca de la apoptosis es inducida por interacciones receptor-ligando. Los ligandos y receptores más importantes para la apoptosis pertenecen a la superfamilia del factor de necrosis tumoral. Los ligandos pueden interactuar con uno o más receptores, y la mayoría de los receptores son proteínas transmembrana con un N-terminal extracelular que interactúa con el ligando y un C-terminal intracelular que tiene un dominio de muerte(88). La interacción con este dominio de muerte puede activar caspasas efectoras a través de varias vías. Por ejemplo, la interacción FAS/FASL junto con las proteínas adaptadoras puede unirse a las procaspasas 8 y 10 y posteriormente activar las caspasas efectoras por autocatálisis(89) o formar complejos proteicos que activan o inhiben las caspasas, como ocurre con el receptor del TNF(90,91). La apoptosis es un mecanismo esencial para mantener la homeostasis celular(92-98) y también representa un mecanismo de defensa en la respuesta inmune, especialmente contra patógenos intracelulares(99).
Papel de la apoptosis en la infección por Mycobacterium bovis
En las infecciones por micobacterias, la apoptosis se ha asociado con una menor diseminación y viabilidad bacteriana(1,2). Sin embargo, cepas y antígenos micobacterianos virulentos pueden inhibir la apoptosis en células previamente infectadas(2,3).
Micobacterias completas
Una de las primeras publicaciones que informó sobre la apoptosis en macrófagos bovinos infectados con M. bovis mostró que la muerte celular se producía tan pronto como 4 h después de la infección utilizando diferentes multiplicidades de infección (MDI). Los autores concluyeron que la apoptosis dependía del tiempo y de la MDI(5). Utilizando el mismo modelo de células, la apoptosis se potenció con IFN-γ/LPS y disminuyó con el bloqueo de TNF-α. Además, en presencia de IL-10, la replicación intracelular micobacteriana se relacionó inversamente con la apoptosis, sugiriendo que la apoptosis desempeña un papel protector contra la infección(6).
El grupo de Rodríguez comparó ratones previamente infectados con cepas atenuadas de M. bovis y con cepas virulentas. Identificaron que la cepa virulenta tenía una mayor capacidad para inhibir la apoptosis en los macrófagos alveolares. Además, la apoptosis fue disminuida por IL-10 e incrementada por TNF-α(7). Los hallazgos anteriores se llevaron a cabo in vivo e in vitro en macrófagos infectados con M. bovis. Este estudio demuestra que las micobacterias modulan la apoptosis a través de la producción de citocinas, el nivel de virulencia y la dosis de exposición. En el Cuadro 1 se presentan algunos de los hallazgos más relevantes de la apoptosis en la infección por el principal agente causal de la TBb.
La resistencia natural contra una enfermedad se define como la capacidad del hospedero para resistir el desarrollo de una enfermedad después de la primera exposición al patógeno y sin inmunización previa(8). La resistencia natural a las infecciones por micobacterias en bovinos ha sido reportada por varios autores. Por ejemplo, el grupo de Esquivel-Solís(9), comparó la apoptosis y la actividad microbicida en macrófagos bovinos resistentes y susceptibles infectados con M. bovis. Los hallazgos indican que la apoptosis aumentó en macrófagos con altos niveles de NO, sugiriendo una relación entre la apoptosis y la actividad microbicida en el fenotipo resistente(9). Estos resultados coinciden con los obtenidos a partir de macrófagos resistentes infectados con M. tuberculosis(100,101). Se estudió el efecto de la IL-4 en macrófagos bovinos en ambos fenotipos. Los resultados muestran una disminución en la expresión de genes proinflamatorios y una menor tendencia a la apoptosis en macrófagos resistentes, evidenciando que la activación alternativa por IL-4 aumentó la susceptibilidad a la infección en macrófagos resistentes(102). También se evaluó la relación entre la producción de NO, la apoptosis y la supervivencia intracelular de micobacterias en células dendríticas de ratones. La apoptosis (fragmentación del ADN y caspasas 3, 6 y 9) y la concentración bacteriana se cuantificaron en ausencia o presencia de un inhibidor de la iNOS. Los resultados de este estudio mostraron que: a) la población infectada con BCG mostró más apoptosis en comparación con M. bovis, b) en presencia del inhibidor, la apoptosis se redujo significativamente en ambas poblaciones infectadas, y c) M. bovis sobrevivió mejor que BCG en CD. Estos resultados sugieren que la reducción de la producción de NO por las células dendríticas debido a la infección por M. bovis modula el desarrollo de la apoptosis e incrementa la posibilidad de supervivencia de las micobacterias(52). Estos resultados destacan el papel del óxido nítrico en la apoptosis en las primeras fases de la infección.
Varios grupos de investigación se han centrado en dianas intracelulares específicas para entender los mecanismos y orgánulos implicados en la apoptosis. Vega et al(103) en 2007 sugirieron una asociación entre la apoptosis y la translocación nuclear del factor inductor de apoptosis (AIF) y la despolarización de la membrana mitocondrial en macrófagos expuestos a un extracto de proteína de M. bovis(103). Esto impulsó la investigación de otros componentes asociados a la apoptosis, por ejemplo, el impacto de la permeabilidad mitocondrial en la fragmentación del ADN y la viabilidad micobacteriana en macrófagos bovinos infectados con M. bovis. La fragmentación del ADN disminuyó independientemente de la actividad de las caspasas cuando se inhibió la permeabilidad mitocondrial. Además, la translocación de AIF y Endonucleasa G (Endo-G) al núcleo, medida por inmunoblot, aumentó 15 y 43 veces, respectivamente, y la viabilidad de las micobacterias intracelulares aumentó en un 26 %(104). Estos resultados apoyan la idea de que la translocación de Endo-G al núcleo también está involucrada en la fragmentación del ADN como resultado de la infección por M. bovis al alterar la permeabilidad mitocondrial. La identificación de moléculas como Endo G y AIF en el núcleo y la disminución de la viabilidad micobacteriana intracelular en ausencia de caspasas activadas, sugieren que la activación de las caspasas no es necesaria para la fragmentación del ADN y revelan la existencia de diferentes mecanismos en el desarrollo y modulación de la apoptosis, especialmente durante la infección.
También se evaluó el estrés mitocondrial inducido por la infección por M. bovis en células THP-1. Se encontró que las caspasas apoptóticas modulan negativamente la producción de IFN-β al reducir la translocación nuclear de p-IRF3 (Factor Regulador de Interferón 3)(105). Esto representa un escenario beneficioso para el hospedero, ya que un menor IFN-β en la infección por M. bovis se ha asociado con un mejor pronóstico(106).
Se investigó el estrés del retículo endoplásmico inducido por infección micobacteriana en macrófagos murinos previamente infectados con M. bovis. Este estudio mostró una mayor supervivencia intracelular de las micobacterias al adicionar un inhibidor del estrés del RE, que moduló directamente el porcentaje de células apoptóticas(107). La relación entre la apoptosis y la funcionalidad de las mitocondrias y del RE y su impacto en la viabilidad de las micobacterias intracelulares destaca el efecto protector de la apoptosis frente a la infección(108). Sin embargo, estos orgánulos y vías también pueden convertirse en objetivos para la modulación de micobacterias.
Las caspasas activadas han sido utilizadas como único marcador de apoptosis en infecciones por micobacterias(109). No obstante, se ha identificado apoptosis independiente de caspasas en bovinos y búfalos infectados con M. bovis(103,110). Además, dado que la apoptosis limita el crecimiento intracelular de las micobacterias, la ausencia de activación de caspasas podría representar un mecanismo de evasión de la muerte celular por M. bovis(107).
La mayoría de los abordajes para investigar la apoptosis en la infección por micobacterias se han llevado a cabo en modelos celulares (principalmente macrófagos), lo que permite estudiar el papel protector de la apoptosis en la fase aguda. Sin embargo, debido a la naturaleza persistente de la infección, la apoptosis también debe estudiarse en la fase crónica de la infección por micobacterias. El grupo de Cherdantseva informó que las células apoptóticas correspondían a aproximadamente el 11 % de las células en granulomas de pulmón de ratones infectados con M. bovis-BCG después de 180 días de infección. Estos datos sugieren que, a pesar de que la apoptosis es inducida a nivel tisular, es insuficiente para eliminar las micobacterias durante el desarrollo del proceso patológico(111).
Proteínas de M. bovis
El doble papel de la apoptosis en la infección por micobacterias se ha observado en estudios con antígenos individuales de M. tuberculosis, en los que los antígenos se clasifican como proapoptóticos o antiapoptóticos. Estos estudios sugieren que las micobacterias modulan el mecanismo de muerte celular a través de la dinámica de la expresión antigénica(2). En este contexto, para identificar proteínas específicas inductoras de apoptosis de M. bovis, se expusieron macrófagos bovinos a diferentes fracciones de extracto de proteínas. La apoptosis independiente de la caspasa fue inducida por dos proteínas recombinantes de M. tuberculosis, hsp70 y hemaglutinina de unión a heparina (HBHA), que tienen alta homología con M. bovis en macrófagos bovinos(112). En este sentido, se han realizado esfuerzos para determinar el perfil proteico en los extractos proteicos de M. bovis. Mediante espectrofotometría de masas, se identificaron MPB70, MPB83 y chaperonina de 60 kDa como las principales proteínas candidatas que inducen la apoptosis independiente de caspasas(28).
M. bovis y M. tuberculosis tienen una alta homología genómica(113). Por lo tanto, investigar el efecto modulador ejercido por proteínas altamente homólogas en las dos especies podría identificar los nuevos mecanismos en la muerte celular. Este conocimiento nos permitiría comprender las particularidades de la infección y la patogenia general de la TBb.
En conjunto, los resultados mencionados anteriormente indican que la apoptosis es un evento multifactorial que involucra características de las bacterias (como la virulencia, el tiempo y la multiplicidad de la infección) y características intrínsecas del hospedero afectado. No obstante, a pesar del efecto de estas múltiples variables, la apoptosis y el crecimiento intracelular de micobacterias en macrófagos bovinos están inversamente correlacionados, lo que sugiere que la apoptosis en la infección por M. bovis representa un mecanismo de defensa del hospedero.
Necrosis o muerte celular accidental
El término necrosis proviene del griego “necro”, que significa cadáver o muerte y “osis”, que significa condición o estado. La necrosis se utilizó para describir la muerte morfológica de las células como resultado de una infección, daño celular, estímulos nocivos o daño mecánico; por lo tanto, se pensó que la necrosis se debía a cambios bruscos que conducían a la muerte celular accidental(114). El diagnóstico anatomopatológico evalúa las características macroscópicas y microscópicas del tejido afectado y clasifica la necrosis en coagulativa, fibrinoide, hemorrágica y caseosa(115).
En la infección por M. bovis, la necrosis está presente en las etapas avanzadas de las lesiones granulomatosas(65). Además, la necrosis no regulada se ha asociado con una mayor diseminación de la infección por micobacterias(116). Un análisis de granulomas de ganado infectado naturalmente con M. bovis mostró grandes áreas necróticas con calcificación central, sin cápsula de tejido conectivo y pocas células gigantes. La necrosis fue la muerte celular predominante observada, y se acompañó de más antígenos micobacterianos, lo que se observó principalmente en terneros(24). Además, la inducción de necrosis con peróxido de hidrógeno en macrófagos infectados por M. tuberculosis favorece el escape de las bacterias al medio extracelular sin afectar su viabilidad(117). Además, otros ensayos de inducción de necrosis permiten la salida y proliferación de micobacterias(118).
Diferentes tipos de muerte celular regulada
En las últimas décadas, los avances bioquímicos y moleculares han permitido descubrir algunos tipos de necrosis que no son accidentales, sino que siguen vías de señalización reguladas que producen una morfología necrótica(99,100,119,120). La descripción de estas vías de señalización ha ayudado a definir las diversas vías de muerte celular que conducen a la muerte celular necrótica. Entre los diferentes tipos de muerte celular necrótica se encuentran la necroptosis, piroptosis, entre otras(99,101,102,103,119,121,122,123).
Piroptosis
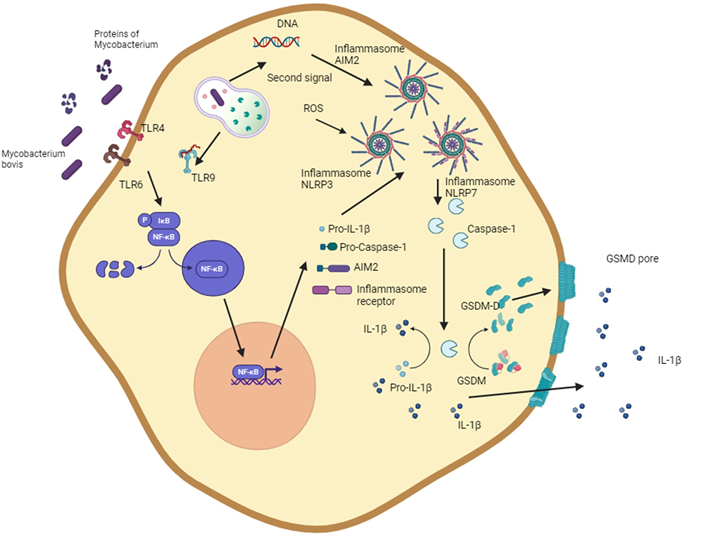
El término piroptosis se deriva del griego “pyro” (fuego, fiebre) y “ptosis” (caída)(104). La piroptosis se describió por primera vez en modelos de infección in vitro por Salmonella y Shigella, en los que la caspasa 1 inició la muerte celular(104,105,106,124,125,126). La piroptosis es una muerte celular inflamatoria caracterizada clásicamente por el inflamasoma, caspasa 1, gasdermina D (GSDMD) y la liberación de IL-1β e IL18 (Figura 1).
Figura 1: Vías de inducción de la piroptosis por Mycobacterium bovis
El diagrama muestra la capacidad de M. bovis y las proteínas micobacterianas para activar los inflamasomas NLRP3 y AIM2. La activación del inflamasoma NLRP3 se inicia a través de receptores de reconocimiento de patrones y luego por múltiples estímulos como la generación de especies reactivas de oxígeno, eflujo de potasio o componentes lisosomales. La activación del inflamasoma AIM2 se inicia mediante el reconocimiento del ADN bacteriano. El ensamblaje del inflamasoma conduce a la maduración de la IL-1β y a la escisión de la gasdermina, formando gasdermina D que daña la membrana celular y provoca la muerte celular necrótica. Esta figura se creó utilizando BioRender.com.
El inflamasoma, que se activa en la piroptosis, consta de estructuras multiproteicas que incluyen un receptor de las familias de receptores NLR (receptores similares al dominio de oligomerización de unión a nucleótidos) o receptores similares al mieloma AIM 2 (AIM2), así como la ASC (proteína similar a una mota asociada a la apoptosis que contiene una CARD) y procaspasa 1(107,108,127,128). Sin embargo, con menos frecuencia, la piroptosis puede activarse por una vía alternativa. La activación del inflamasoma conduce a la activación de caspasas inflamatorias (caspasas-1,-4,-5 en humanos y caspasas-1 y -11 en ratones) y a la escisión de la familia de las interleucinas-1 y GSDMD. La GSDMD activa puede ensamblarse para formar poros en la membrana celular y generar un desequilibrio osmótico que conduce a la muerte celular en un ambiente inflamatorio(109,110,129,130).
M. bovis puede inducir piroptosis en células de macrófagos y líneas celulares derivadas de macrófagos (Cuadro 1). La cepa de la bacteria, la multiplicidad de la infección y el tiempo después de la infección se encuentran entre los factores que favorecen la piroptosis(10,131,132,133). Los principales mecanismos que inducen la piroptosis están relacionados con la activación canónica de los inflamasomas (Figura 1). NLRP7, que reconoce glicoproteínas bacterianas; AIM2, que reconoce el ADN bicatenario; y NLRP3, que es activada por diversas señales, como eflujo de potasio, ERO, ATP extracelular, toxinas formadoras de poros y media la piroptosis asociada a la infección por M. bovis(111,112,113,131,132,133). La activación de los inflamasomas afecta la producción de IL-1β, IL-18 e IL-33, generando un ambiente inflamatorio que ayuda a controlar la infección producida por micobacterias(132).
La activación del inflamasoma NLRP3 requiere dos señales y genera un entorno inflamatorio. La estimulación de macrófagos con LPS aumenta la IL-1 β y el óxido nítrico, lo que puede limitar el crecimiento intracelular de las micobacterias(9,10). La activación del inflamasoma por macrófagos bovinos infectados con M. bovis disminuye el crecimiento intracelular de micobacterias(10).
El ambiente inflamatorio generado por la piroptosis puede regular la proliferación de bacterias, reclutando células inmunitarias que ayudan a controlar las infecciones bacterianas. No obstante, la piroptosis puede causar daño tisular, por lo tanto, representa un fuerte mecanismo que tienen algunas células hospederas para controlar el crecimiento intracelular bacteriano. Cabe destacar que actualmente no hay información sobre qué cepas de bacterias suelen inducir la piroptosis. También se desconoce si el crecimiento bacteriano es controlado o si algunas bacterias inducen este tipo de muerte celular para escapar de las células e infectar los tejidos circundantes(12,134).
Autofagia
El término autofagia se deriva del griego “auto” (yo) y “phagen” (comer). La autofagia es una vía altamente conservada que degrada los componentes celulares utilizando lisosomas(135). La autofagia es un mecanismo regulado que permite a las células sobrevivir bajo privación de nutrientes o condiciones adversas. Sin embargo, la autofagia también puede causar la muerte celular (muerte celular dependiente de la autofagia). Este mecanismo puede ocurrir concomitantemente con otro tipo de muerte celular, como la apoptosis, o comenzar como autofagia y desencadenar la apoptosis(136).
Se ha demostrado que la autofagia limita las bacterias intracelulares. Algunas de las moléculas involucradas en este proceso son la proteína relacionada con mieloides 8/14 y la proteína inducible por interferón-γ 204 (IFI204) que induce la autofagia en células mononucleares de sangre periférica y células THP1 de manera dependiente de ERO, lo que inhibe el crecimiento intracelular de Mycobacterium BCG (Cuadro 1)(137). Además, IFI204 es un sensor de ADN que activa la respuesta inmunitaria innata, incluida la autofagia y la producción de interferón-β (IFN-β). Las proteínas IFI204 están implicadas en las respuestas del IFN-β mediante el reclutamiento de STING para activar las vías TBK-1-IRF3. La inducción de la autofagia por IFI204 induce la fosforilación de TBK-1 para inhibir la supervivencia de M. bovis en macrófagos(138).
Es importante destacar que M. bovis puede evadir la autofagia. Uno de los mecanismos consiste en la inhibición específica de la autofagia responsable del control de los organismos intracelulares (xenofagia), por ejemplo, a través de la activación de la vía indicadora PINK1-PRKN/Parkin involucrada en la mitofagia, lo que genera una competencia de ambas vías por p-TBK1 que conduce a una disminución de la xenofagia y a la supervivencia de las micobacterias(139). Se evaluó el papel del microARN miR-199a en macrófagos derivados de la médula ósea, el pulmón y el bazo de ratones infectados con M. bovis. La infección aumentó la expresión de miR-199a, y esto suprimió la autofagia al bloquear la maduración de los fagolisosomas a través de la interacción con la quinasa 1 de unión a TANK. Estos cambios condujeron a un aumento de la supervivencia intracelular de las micobacterias. Estos resultados proporcionan un mecanismo para que M. bovis evada la eliminación(137).
Aunque el desarrollo de la autofagia participa en el mantenimiento del equilibrio celular, también puede funcionar como un mecanismo de respuesta inmunitaria innata que limita el crecimiento de las bacterias intracelulares. En las infecciones por M. bovis, la autofagia es inducida por bacterias de baja virulencia, lo que sugiere que M. bovis también puede modificar los procesos involucrados en el mantenimiento de la homeostasis celular(140,141).
Conclusiones
Independientemente de la influencia de diferentes variables (como la virulencia, el tiempo, la especie y el fenotipo de resistencia del hospedero) en la apoptosis, los resultados experimentales sugieren que la muerte celular por apoptosis ayuda a controlar el crecimiento bacteriano.
El efecto inhibidor bacteriano sobre la apoptosis, la redirección de la autofagia y la inducción de la muerte celular inflamatoria, como la necrosis y la piroptosis, pueden ser mecanismos bacterianos para evadir la respuesta inmunitaria del hospedero.
Aunque las condiciones experimentales permiten la detección de un tipo específico de muerte celular, también se ha observado la activación simultánea de múltiples tipos de muerte celular, conocida como panoptosis, en la infección por M. tuberculosis. Este escenario abre la posibilidad de estudiar la infección por M. bovis de una manera global que considere todas las variables experimentales y fases de los diferentes tipos de muerte celular.
La alta adaptabilidad de M. bovis y el papel clave de la muerte celular en la activación inmunitaria ponen de manifiesto la necesidad de realizar más estudios sobre la muerte celular regulada y no regulada. Estos estudios aumentarán la comprensión de la infección bovina y ayudarán a desarrollar nuevas estrategias para contrarrestar la tuberculosis bovina.
Los puntos más importantes de esta revisión se pueden numerar en: 1) La muerte celular por apoptosis ayuda a controlar el crecimiento bacteriano. 2) La autofagia es un mecanismo conservado que limita la replicación intracelular de las micobacterias. 3) La piroptosis es un mecanismo extremo que ayuda a controlar M. bovis a costa de dañar el tejido del hospedero. 4) La necrosis permitirá el escape y proliferación de micobacterias.
Agradecimientos
Agradecemos la beca posdoctoral de la DGAPA UNAM para Paola Andrea Ortega Portilla como beneficiaria. Omar Escobar-Chavarría recibió apoyo de una beca del Consejo Nacional de Humanidades, Ciencia y Tecnología CONAHCYT. Esta investigación fue financiada con una beca de la Universidad Nacional Autónoma de México, DGAPA-PAPIIT IG201521 y DGAPA-PAPIIT IG200918.
Declaración de conflicto de intereses
Los autores declaran que no tienen intereses financieros o relaciones personales contrapuestas que puedan haber influido en el trabajo reportado en este trabajo.
Cuadro 1: Muerte celular en la infección por Mycobacterium bovis
+Molécula diana: molécula seleccionada para evaluar la muerte celular, (R): Fenotipo de resistencia, ND: No determinado, Carga micobacteriana*: Cuantificada en presencia del tipo específico de muerte concomitantemente, AIF: Factor inductor de apoptosis, HBHA: hemaglutinina de unión a heparina, AIM2: ausente en el melanoma 2, LDH: lactato deshidrogenasa, NLRP3: NOD, repeticiones ricas en leucina y proteína 3 que contiene dominio de pirina, NOD, repeticiones ricas en leucina y proteína 7 que contiene dominio de pirina, LC3: Proteína 1A/1B de cadena ligera 3 asociada a microtúbulos, LAMP-1: proteína de membrana asociada a lisosomas, HSPD1: proteína 1 de choque térmico de 60 kDa, PPE: proteínas de familias que contienen motivos Pro-Glu (PE) y Pro-Pro-Glu (PPE), BMDM: macrófagos derivados de la médula ósea, MRP8/14: proteínas relacionadas con la mieloides (MRP) 8 y 14, LRG-47: proteína inducible por IFN Irgm1.
Literatura citada: