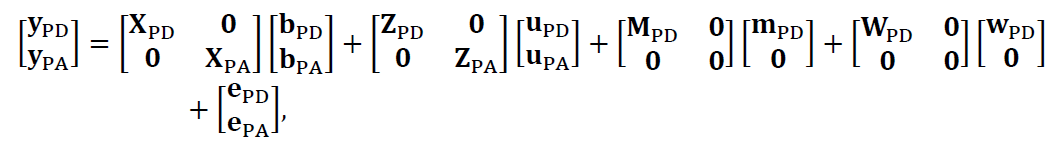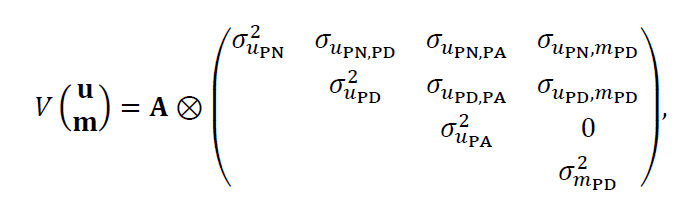https://doi.org/10.22319/rmcp.v16i1.6648
Artículo
Optimización de la predicción del valor de reproducción en ganado Braunvieh por medio del análisis multirasgo de rasgos de crecimiento
Mohammad Ali Nilforooshan a
Agustín Ruíz-Flores b
Paulino Pérez-Rodríguez c
Thierry Pabiou d
Luis Antonio Saavedra-Jiménez e
Jonathan Emanuel Valerio-Hernández f*
a Livestock Improvement Corporation, Research & Development Division, Hamilton 3240, Waikato, New Zealand.
b Universidad Autónoma Chapingo. Posgrado en Producción Animal. Estado de México, México.
c Colegio de Postgraduados. Socio Economía Estadística e Informática. Estado de México, México.
d The Irish Cattle Breeding Federation. Link road, Ballingcollig, Co. Cork, Ireland.
e Universidad Autónoma de Guerrero. Facultad de Medicina Veterinaria y Zootecnia No. 2. Guerrero, México.
f Universidad Nacional Autónoma de México. Facultad de Medicina Veterinaria y Zootecnia. Ciudad de México, México.
*Autor de correspondencia: jevh_93@hotmail.com
Resumen:
Actualmente, la evaluación genética de los rasgos de crecimiento (peso al nacer - PN, peso al destete - PD y peso al año - PA) en el ganado Braunvieh en México se lleva a cabo por medio de modelos univariados para el PN y bivariados para el PD y PA. La precisión de las evaluaciones genéticas se puede mejorar mediante un modelo trivariado. Se analizó tanto el sesgo en las evaluaciones univariadas y bivariadas causado por los rasgos faltantes en el análisis, como el aumento en la precisión producido por el análisis trivariado. Los datos de pedigrí y de rendimiento se obtuvieron de la Asociación Mexicana de Criadores de Ganado Suizo de Registro. Después de editar los datos, se realizaron análisis univariados, bivariados y trivariados y se hicieron comparaciones entre ellos. Se empleó una estrategia simple de reducción del conjunto de datos, reduciendo notablemente la cantidad de datos en los análisis. Los animales excluidos de los análisis se evaluaron a partir de las soluciones de los animales incluidos en los análisis, lo cual requirió mucho menos poder de computación. El análisis bivariado mostró evaluaciones sesgadas del PD y PA y de las tendencias genéticas. Las tendencias genéticas se subestimaron en los animales jóvenes. Desde mediados de la década de 1990, todos los rasgos mostraron una ganancia genética constante. El sesgo se debió a la preselección natural/artificial para PN. La inclusión del PN en el análisis trivariado ayudó a tomar en cuenta la información de preselección. La evaluación univariada del PN y la tendencia genética fueron imparciales. También, el PN aumentó menos en precisión a partir del PD y PA que el PD y PA a partir del PN. Se recomienda el análisis trivariado de los rasgos con una reducción del conjunto de datos para reducir el poder computacional requerido para procesar los datos.
Palabras clave: Precisión, Modelo animal, Sesgo, Valor de reproducción, Braunvieh, Multirasgo, Preselección, Reducción, Univariado.
Recibido: 20/02/2024
Aceptado: 07/11/2024
Introducción
El ganado Pardo Suizo Europeo se introdujo a México a mediados del siglo XIX(1,2). En 1967 se creó la Asociación Mexicana de Criadores de Ganado Suizo de Registro (AMCGSR), y en 1968 se registraron las variantes lechera (Pardo Suizo Americano) y de doble propósito (Pardo Suizo Europeo o Braunvieh)(3). Las potencialidades de la raza, como buena fertilidad, rusticidad, adaptabilidad y producción lechera y de carne, han posicionado a esta raza como una de las favoritas para cruzar con la raza Cebú (Bos indicus) en los sistemas de producción en el trópico mexicano. Durante las últimas dos décadas, el ganado Braunvieh mexicano ha ido desplazando de manera gradual a B. indicus(4).
La primera evaluación genética nacional de la raza Braunvieh se realizó en 2003 por la AMCGSR, y desde aquel entonces la asociación hace evaluaciones periódicas y seguimiento de tendencias genéticas en esta raza(5). Actualmente, estas evaluaciones involucran el peso al nacimiento (PN), el peso al destete (PD), el peso al año de edad (PA) y la circunferencia escrotal (CE); el modelo para la raza Pardo Suizo americano también incluye el volumen de leche ajustado a 210 días de lactancia(6). La reproducción de Braunvieh ha permitido ganancias genéticas; sin embargo, los niveles de ganancia genética no han sido tantos como se esperaba. Larios-Sarabia et al(7) reportaron disminuciones en la tendencia genética de la producción de leche en hatos Jersey y Pardo Suizo americano en México, en parte debido a que los objetivos de selección han diferido entre los hatos. Estos autores plantearon la necesidad de revisar y reestructurar los programas nacionales de mejoramiento genético para esas poblaciones lecheras.
Las evaluaciones genéticas que incluyen múltiples rasgos (multirasgo) han sido muy útiles en el mejoramiento genético de los animales(8,9). Este tipo de modelo tiene varias ventajas en comparación con los modelos univariados: una mayor magnitud de las heredabilidades estimadas(10); reducción del sesgo introducido por la selección secuencial(8,11); un aumento en la precisión de los valores de reproducción(11,12); y mejores propiedades de los estimadores, especialmente para datos incompletos(9). Además, los modelos multirasgo aprovechan mejor la información disponible por medio de correlaciones entre los rasgos, y desde luego, producen evaluaciones más precisas(11). Sin embargo, las evaluaciones multirasgo tienen un mayor requerimiento de poder computacional, que genera mayores costos, para construir el sistema de ecuaciones y resolver un gran conjunto de ecuaciones. Además, su convergencia es más lenta (es decir, requiere de más interacciones que son más lentas) debido a un mayor número de elementos distintos a cero en la matriz de coeficientes que se ubican fuera de la diagonal(11).
En los modelos multirasgo, los fenotipos de un rasgo sirven como fenotipos (con una importancia ponderada) para los demás rasgos. Esto genera más valores de reproducción, como los valores de reproducción para rasgos medidos más tarde en la vida. Por ejemplo, un ternero recién nacido recibirá valores de reproducción para PD y PA en función de su propio fenotipo de PN y de los fenotipos de sus parientes para cualquiera de estos tres rasgos. Esos valores de crianza son más precisos que los promedios de los padres del ternero (valor de crianza) para PD y PA.
La AMCGSR evalúa de manera periódica los rasgos de PN, PD, PA y CE, y comunica los resultados de la evaluación genética a las partes interesadas y a los ganaderos. La asociación tiene como objetivo aumentar la productividad del ganado, proteger los intereses de los criadores y promover la raza. Debido a la gran cantidad de datos históricos disponibles y a un poder de computación limitado, por muchos años se han evaluado a estos rasgos por separado usando modelos animales univariados. Desde el 2016, se han evaluado los rasgos de PD y PA con un modelo animal bivariado. Para seguir mejorando las evaluaciones, el siguiente paso podría ser evaluar los tres rasgos de crecimiento en conjunto usando un solo modelo animal multirasgo. De esa manera se liberaría a las evaluaciones de PD y PA de la preselección (natural/artificial) para el PN, y los tres rasgos se beneficiarían de la información adicional en el análisis.
En un esfuerzo para desarrollar un análisis genético a este nivel para la raza Braunvieh, el presente estudio se enfocó en tres objetivos. El primero es desarrollar un modelo multirasgo para la evaluación conjunta del PN, PD y PA en la población mexicana de la raza Braunvieh en un entorno computacional restringido. El segundo es cuantificar el sesgo en las evaluaciones genéticas actuales para los rasgos de crecimiento (es decir, la evaluación univariada de PN y la evaluación bivariada de PD-PA). El tercero es incrementar la precisión de la evaluación al analizar los tres rasgos de crecimiento de manera simultánea.
Material y métodos
Datos
Los datos de pedigrí y rendimiento se obtuvieron de la AMCGSR. Para cada rasgo (PN, PD y PA), el criterio de inclusión de datos fue que un hato tuviera un mínimo de cuatro registros de rendimiento. Se eliminaron los registros de animales nacidos por transferencia de embriones (debido a la falta de identificación de las vacas receptoras) o en los cuales no se identificaron a ambos padres. Los fenotipos del PD se limitaron a aquellos tomados en un rango de 195 y 285 días de edad, y luego se ajustaron a un objetivo de 240 días de edad. Los fenotipos del PA se limitaron a aquellos tomados en un rango de 320 y 410 días de edad, y luego se ajustaron a un objetivo de 365 días de edad. Luego, se descartaron los registros fuera del rango del rasgo de media ± 3 DE. Además, se descartaron los fenotipos si la edad de la madre al nacimiento del animal estaba fuera del rango de 20 a 180 meses. Los grupos contemporáneos se definieron dentro de un rasgo por el rebaño (256), el año (1901-2020) y la estación (lluviosa o seca) de la ponderación. Se requirió que los grupos contemporáneos tuvieran un tamaño mínimo de tres animales. Después de descartar los grupos contemporáneos que no cumplieron con el tamaño mínimo, quedaron 2,532 grupos contemporáneos que representaron 37,738 animales.
Reducción del conjunto de datos
Hasta el 2020, había registros de un total de 193,442 animales en el pedigrí. Para la reducción del conjunto de datos se aplicó una sencilla estrategia de la extracción ascendente del pedigrí de los animales con un fenotipo en al menos uno de los tres rasgos (37,738). El subconjunto del pedigrí extraído de esos 37,738 animales contenía 64,501 animales nacidos entre 1950 y 2020. Se esperaba que esta reducción en el universo reduciría el tiempo y el poder de computación necesarios para producir los resultados. Los animales excluidos de los análisis no tenían contribución al fenotipo, pero sus valores de reproducción (valores de reproducción excluidos – VRE) y la precisión de estos (r) se estimaron de manera iterativa a partir de los datos de sus padres.
(VREprogenie = (VREpadre + VREmadre)/2 y rprogenie=0.5×rpadre2+rmadre2)
El procedimiento iterativo fue el siguiente:
Calcular los valores de reproducción y sus precisiones para los animales los padres de los cuales (si había registros) se encuentran en subconjunto, basado en los valores de los padres y su precisión.
Agregar las filas de pedigrí para estos animales al subconjunto del pedigrí.
Repetir los pasos 1 y 2 mientras quedaban animales para agregar al subconjunto del pedigrí.
El subconjunto de pedigrí incluyó 21,405 machos, 43,096 hembras, 3,321 padres y 29,700 madres. Se calcularon los valores de reproducción y su precisión de 171,390 animales utilizando el procedimiento iterativo anterior. Los 22,052 animales restantes se encontraban en árboles de pedigrí que no recibieron ninguna contribución fenotípica, los cuales no se consideraron en el análisis. Independientemente del análisis, se asignaron a estos animales un valor de reproducción y una precisión de 0.
Se llevaron a cabo los análisis con un servidor t2.medium AWS (Amazon Web Services) Ubuntu 20.04 LTS con dos CPU y 4 GB de RAM.
Análisis
Tal como se hace actualmente en la AMCGSR, se analizó el PN con un modelo univariado, y los PD y PA se analizaron con uno bivariado:
yPN=XPNbPN+ZPNuPN+ePN,
donde y, b, u, m, w, y e son los vectores de los fenotipos, los efectos fijos, el efecto genético directo, el efecto genético maternal, el efecto ambiental permanente maternal, y los residuales, respectivamente. Los matrices X y Z vinculan los fenotipos a los efectos fijos y a los animales, respectivamente. Los matrices M y W vinculan los fenotipos a las madres. Para analizar los tres rasgos un conjunto, se aplicó un modelo trivariado:
Las estructuras del componente de varianza fueron:
V(e) = I⊗R, y V(wPD) = Iw2. A es la matriz de relaciones del numerador (basado en el pedigrí), y R es la matriz de la covarianza del residual rasgo × rasgo.
Se incluyeron en los modelos los efectos fijos de clase de sexo (todos los rasgos), el régimen de alimentación con leche (3 niveles – solo PD), el régimen de alimentación post-destete (3 niveles – solo PA), y el grupo contemporáneo (todos los rasgos). Además, se incluyeron los efectos fijos de regresión de la edad de la madre al momento del nacimiento (emn), emn2, y el porcentaje de pureza de Braunvieh. Se describen estos efectos en un estudio previo(13). La covariable de pureza tuvo un mínimo de 0.880, una media de 0.996 y una mediana de 1. La fila del 10.7% tuvo una pureza menor a 0.99, y la de 1.9% tuvo una menor a 0.95.
La familia de programas BLUPF90(14) se utilizó para el análisis de datos, incluida la estimación de componentes de varianza utilizando la expectativa-maximización para calcular estimaciones REML (EM-REML) con aceleración, el valor genético, y la predicción de precisión. La construcción de ecuaciones de modelos mixtos y la teoría de modelos animales multirasgo están bien explicadas por Schaeffer(11).
Resultados y discusión
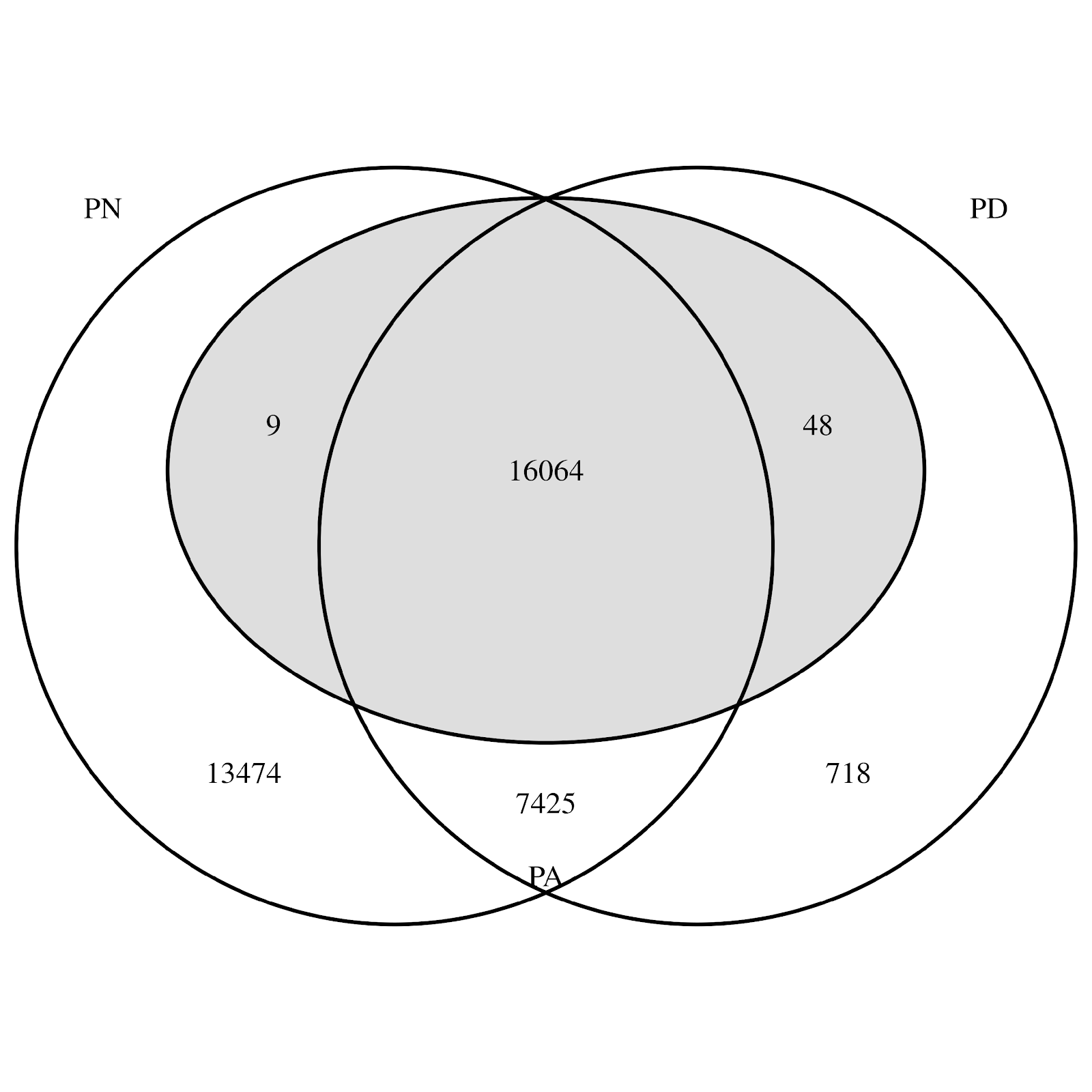
Entre los 37,738 animales fenotipados incluidos en los análisis, 16,064 tenían fenotipos para todos los rasgos y 13,474 tenían fenotipos solo para el PN (Figura 1). De los 16,121 animales fenotipados para el PA, 48 carecían del fenotipo de PN, 9 carecían del fenotipo de PD y ninguno carecía de PN y PD (Figura 1, Cuadro 1).
Figura 1: El número de animales con fenotipos disponibles en combinaciones diferentes de rasgos
PN= peso al nacer (círculo izquierdo); PD= peso al destete (círculo derecho); PA= peso al año de edad (ovalo gris en medio).
Cuadro 1: Estadísticos descriptivos de los fenotipos utilizados en el estudio
PN= peso al nacer; PD= peso al destete; PA= peso al año de edad.
Se estimaron las heredabilidades como h2=u2u2+m2+w2+e2, donde u2, m2, w2, y e2 son los componentes de varianza estimados vinculados a los vectores u, m, w, y e, respectivamente. Faltando cualquier de estos efectos en el modelo, la varianza correspondiente es igual a 0. Las estimaciones de la heredabilidad generadas con el modelo trivariado eran similares a las de los modelos univariado y bivariado (Cuadro 2); por ejemplo, para el PD, hm2=m2u2+m2+w2+e2 disminuyó de 0.021 ± 0.009 en el bivariado a 0.020 ± 0.008 en el trivariado. Desde los análisis univariado/bivariado al análisis trivariado, las covarianzas genéticas cambiaron de
a
y las covarianzas residuales cambiaron de
a
Donde PDm es el efecto genético materno de PD. Los cambios en las correlaciones genéticas y residuales eran mínimas. La varianza ambiental permanente materna de PD cambió de 7.34 en el modelo bivariado a 6.84 en el trivariado.
Cuadro 2: Las heredabilidades generadas por la técnica actual de la AMCGSR, y del modelo trivariado con los tres rasgos de peso
1 Univariado para el PN, y bivariado para los PD y PA.
PN= peso al nacer; PD= peso al destete; PA= peso al año de edad.
A diferencia de los modelos univariados y bivariados, el modelo trivariado incluyó las correlaciones genéticas y residuales entre el PN y los otros dos rasgos. Se basa en el supuesto de que los fenotipos de PN pueden contribuir a mejores evaluaciones genéticas del PD y PA, y viceversa. Además, si hay selección natural o artificial sobre el PN, el modelo trivariado puede eliminar ese sesgo de preselección de las evaluaciones genéticas del PD y PA. Otra característica importante del modelo trivariado es que proporciona estimaciones iniciales de los méritos genéticos de PD y PA para terneros recién nacidos.
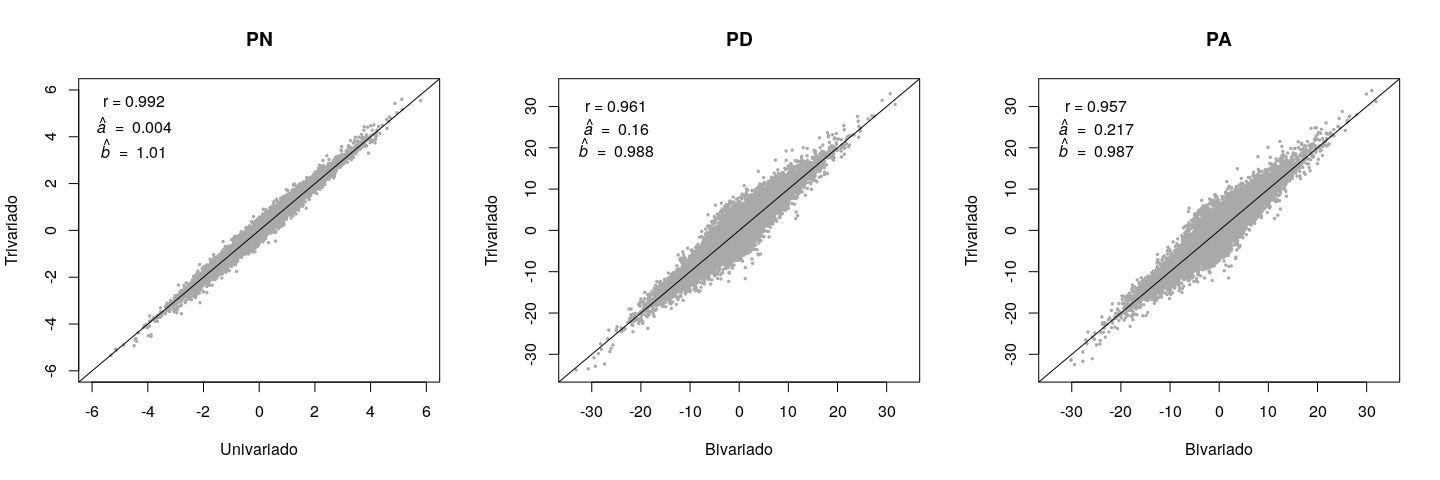
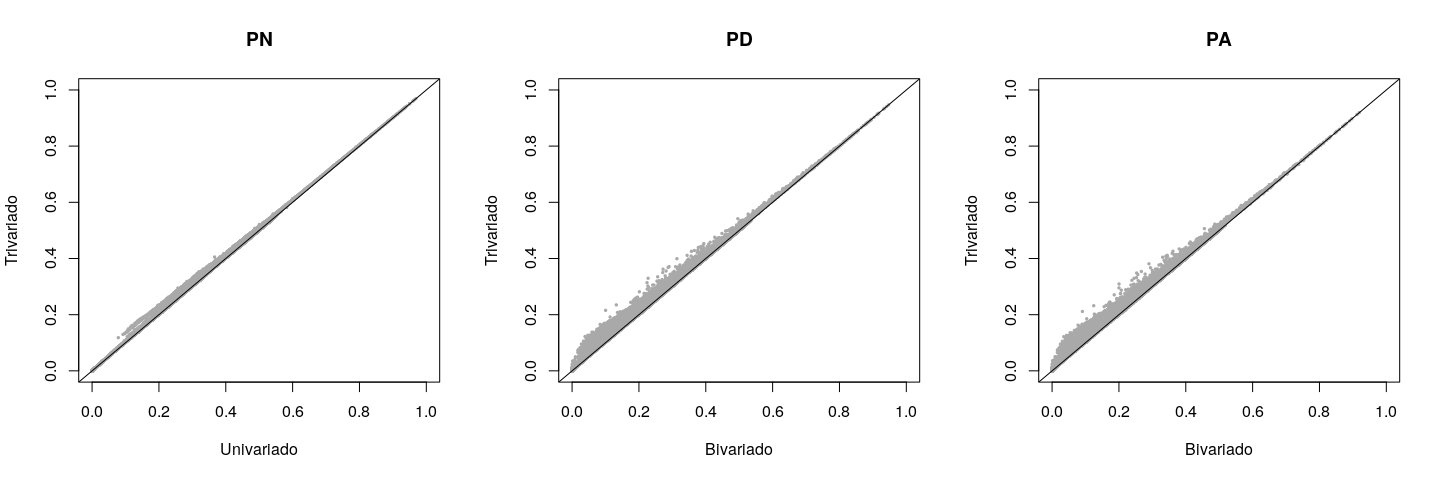
Los valores de reproducción para diferentes rasgos en los modelos univariados/bivariados comparados con los del trivariado se muestran en un diagrama de dispersión que incluye los coeficientes de correlación y regresión correspondientes a cada rasgo (Figura 2). Peso al año de edad (PA) tuvo el coeficiente de correlación más bajo (r= 0.957), la desviación del intercepto de 0 más alto (a= 0.217), y la mayor desviación del coeficiente de regresión (pendiente de la línea de regresión) de 1 (b= 0.987). En contraste, el PN mostró las menores desviaciones. Si el coeficiente de regresión se desvía de 1, hay sesgo, tomando en cuenta que el supuesto verdadero de que los fenotipos PN no introducen sesgo en las evaluaciones del PD y PA. En el análisis bivariado, el sesgo para el PD y PA fue menor y ascendente (b< 1), y se debió al efecto de la preselección natural/artificial sobre el PN.
Figura 2: Comparación entre los valores genéticos estimados por los modelos univariado (PN) o bivariado (PD and PA) y los estimados por el modelo trivariado (PN, PD, y PA)
PN= peso al nacer; PD= peso al destete; PA= peso al año de edad.
La tasa de cambios en el valor genético del PN fue mayor a la esperada debido a la mayor heredabilidad de este rasgo. Sin embargo, los valores genéticos del PN fueron los menos afectados por el análisis trivariado (Figura 2). Probablemente, esto se debió a la ausencia de un efecto de preselección del PD y PA sobre el PN. Aunque la heredabilidad de PA fue menor que la de PD (Cuadro 2), la tasa de cambios en el valor genético (es decir, correlación) fue similar entre los dos rasgos. Esto ocurrió porque la información del PN, por medio del PD, influye directa e indirectamente en la evaluación del PA. En un estudio similar, pero con la raza Angus, se recomienda un modelo univariado para el PN y uno bivariado para el PD y PA(8). Para la raza Tropicarne, el mismo estudio recomienda modelos bivariados tanto para el PN y PD, como para el PD y PA.
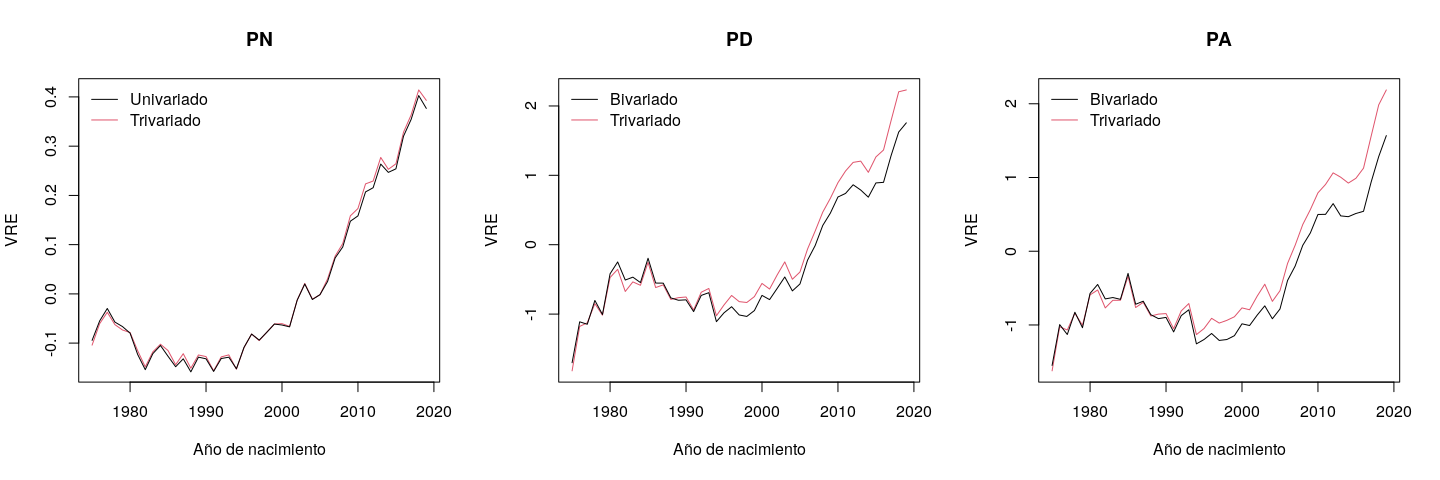
Las tendencias genéticas se analizaron usando los tres rasgos y los tres tipos de modelo (univariado, bivariado y trivariado) con datos de 1975 a 2019 (no se usaron los de 2020 porque solo nacieron 20 animales) (Figura 3). Para todos los rasgos, la tasa de ganancia genética fue lenta o negativa en la década de 1980 al 1990, pero aceleró a partir de mediados de la década de 1990. La tendencia genética de PN no varió entre los modelos univariados y trivariados, apoyando el hallazgo que las evaluaciones univariadas de PN son imparciales (b= 1.01 para PN, Figura 1). Sin embargo, al subestimar los animales jóvenes el análisis bivariado sí mostró sesgo en las tendencias genéticas del PD y PA. Las desviaciones entre los promedios del valor genético fueron mayores para los animales nacidos en los últimos años (Figura 3). Una vez más, esto demuestra la importancia de los datos de PN para evitar/reducir el sesgo debido a la preselección de PN para la evaluación del PD y PA.
Figura 3: Las tendencias genéticas estimadas por los modelos univariado (PN), bivariado (PD and PA), y trivariado (PN, PD, and PA)
PN= peso al nacer; PD= peso al destete; PA= peso al año de edad.
En el análisis del aumento en la precisión desde los modelos univariado (PN) y bivariado (PD y PA) al modelo trivariado, los promedios fueron 0.006 para el PN, 0.010 para PD, y 0.011 para el PA (Figura 4). Existen dos posibles razones para el bajo aumento en la precisión de PN. El primero, es que solo hubo 766 animales con el fenotipo PD y sin el fenotipo PN, de los cuales 48 también tuvieron el fenotipo PA (Figura 1); y el segundo, es que las heredabilidades del PD y PA fueron menores que la heredabilidad del PN (Cuadro 2). El aumento promedio en la precisión de PN para esos 766 animales con el fenotipo PD y sin el de PN fue de 0.028. Sin embargo, se debe mencionar que el aumento en la precisión no se trata solo del fenotipo propio sino también de la contribución del fenotipo de todos los parientes (ponderada por el coeficiente de relación y la heredabilidad). El PD y el PA fueron más precisos a partir de la presencia de PN en el análisis trivariado. Es probable que eso se deba a dos factores: 1) una mayor heredabilidad de PN y; 2) una cantidad grande de animales con el fenotipo PN y sin los fenotipos PD y PA (Figura 1; es decir, los animales con poca contribución de los fenotípicos para el PD y PA reciben contribución fenotípica del rasgo correlacionado de PN). De hecho, los animales con baja precisión la vieron aumentar por medio del análisis trivariado (Figura 4). Esos animales tuvieron una baja contribución fenotípica (probablemente sin fenotipo propio) en el análisis univariado (PN) o bivariado (PD y PA), pero recibieron más contribución fenotípica (a través de su propio desempeño y/o los de sus parientes) por medio del(los) rasgo(s) adicional(es) en el análisis trivariado. Por ejemplo, el aumento en la precisión promedio de PD para los 13,483 animales con PN y sin fenotipos PD fue de 0.037. De manera similar, el aumento en la precisión promedio de PA para los 13,474 animales con PN y sin los fenotipos del PD y PA fue de 0.040. El aumento en la precisión promedio de PA para los 718 animales con los fenotipos del PD y sin los de PN y PA fue de 0.004, y para los 7,425 animales con los fenotipos del PN y PD y sin el de PA fue de 0.015. Los valores genéticos con baja precisión tienden a retroceder hacia la solución de los fundadores, que es 0.
Figura 4: Los valores genéticos estimados por medio de análisis univariado (PN) o bivariado (PD and PA) comparados con los estimados por medio del análisis trivariado (PN, PD, and PA)
PN= peso al nacer; PD= peso al destete; PA= peso al año de edad.
Si la heredabilidad de los rasgos y las correlaciones absolutas entre ellos tienen valores altos, la precisión aumenta. Además, se espera que los resultados para los animales que le falten fenotipos aumenten más en precisión con los rasgos de fenotipos correlacionados. Un estudio de las relaciones entre la heredabilidad, la correlación genética (rg), y la correlación residual (re) entre rasgos usando un modelo animal multirasgo reporta tres principales hallazgos: 1) lo más alto que es el |rg – re|, mayor es el aumento en la precisión; 2) en |rg| < |re|, el rasgo con menos heredabilidad experimenta un mayor aumento en la precisión; y 3) en |rg| > |re|, el rasgo con más heredabilidad experimenta un mayor aumento en la precisión(15).
Conclusiones e implicaciones
Debido a los recursos limitados de computación, por muchos años las evaluaciones genéticas de los rasgos de crecimiento (PN, PD y PA) en el ganado Braunvieh en México se llevaron a cabo con modelos univariados. Solo desde el 2016, el PD y PA se han evaluado con un modelo bivariado. Las evaluaciones de rasgos múltiples son preferibles porque representan un uso más completo de los datos disponibles, y proveen un aumento en la precisión de los resultados a través de las correlaciones entre los rasgos y la eliminación/reducción del sesgo causado por la selección(11). Esta última ventaja es más evidente en los rasgos medidos y seleccionados de manera secuencial. En los resultados, las evaluaciones univariadas de PN fueron imparciales mientras que las evaluaciones bivariadas de PD y PA resultaron ligeramente sesgadas a raíz de la selección (no aleatoria) en PN. Se supone que la selección artificial en PN es débil e involucra un efecto de la selección sobre otros rasgos previos al destete (correlacionados con PN), como la ganancia diaria previa al destete y la selección natural en PN. Lo ideal para corregir dicho sesgo es aplicar un modelo multirasgo que incluya los tres rasgos; se recomienda ampliamente su implementación a la AMCGSR. El incremento del costo computacional que representa el uso del modelo trivariado se puede mitigar aplicando una estrategia de reducción del conjunto de datos, lo cual reduce de manera notable el poder de computación requerido. A los animales descartados como parte de la reducción se pueden evaluar a un bajo costo computacional utilizando las soluciones de los animales en el análisis.
Literatura citada: