https://doi.org/10.22319/rmcp.v15i2.6626
Technical note
Prevalence and infection intensity of honey bee (Apis mellifera) viral diseases in six regions of the state of Jalisco, Mexico
Ana Karen Ramos-Cuellar a
Álvaro De la Mora b
Francisca Contreras-Escareño c
Nuria Morfin d
José María Tapia-González e
José Octavio Macías-Macías e
Tatiana Petukhova f*
Adriana Correa-Benítez a
Ernesto Guzman-Novoa b
a Universidad Nacional Autónoma de México. FMVZ, Departamento de Medicina y Zootecnia de Abejas. Ciudad de México, México.
b University of Guelph. School of Environmental Sciences, 50 Stone Road East, Guelph, ON, N1G 2W1, Canadá.
c Universidad de Guadalajara. CUCSur, Depto. Prod. Agríc., Autlán, Jal., México.
d University of British Columbia. Dept. Biochem. Mol. Biol., Vancouver, BC, Canadá.
e Universidad de Guadalajara. CUSur, Depto. Cienc. Natur., Cd. Guzmán, Jal., México.
f University of Guelph. Department of Population Medicine, Guelph, ON, Canadá.
*Corresponding author: tpetukho@uoguelph.ca
Abstract:
Jalisco is one of the foremost honey-producing states in Mexico. However, there is no information on viral diseases that affect honey bees (Apis mellifera) in the different beekeeping regions of the state. The objective of this study was to determine the prevalence and intensity of four viral diseases of Apis mellifera during the spring, in six regions of Jalisco. Bee samples from 79 colonies were analyzed, of which, 66 % and 38 % were positive for black queen cell virus (BQCV) and deformed wing virus (DWV), respectively. Two viral diseases were not detected, those caused by the Israeli acute paralysis virus (IAPV) and the chronic bee paralysis virus (CBPV). The infection levels of BQCV were relatively low but elevated for DWV, with infection intensities 8,000 higher than those of BQCV. The prevalence of DWV was significantly higher in the regions of the Highlands, Center, and South, while for BQCV there were no differences between regions. For infection intensity, there were no differences between regions for DWV, but there were for BQCV. The regions with the highest infection levels were the South and Center. Surveys during other seasons of the year are recommended to identify possible seasonal viral effects on the bees and to design control strategies.
Keywords: Apis mellifera, Deformed wing virus, Black queen cell virus.
Received: 15/10/2023
Accepted: 23/02/2024
Viral diseases of honey bees (Apis mellifera) are increasingly being associated with colony losses(1), so it is important to know their prevalence and distribution to be able to control them. More than 20 viruses are known to infect honey bees, but few of them appear to have a serious impact on their health. Among them, we can mention deformed wing virus (DWV), black queen cell virus (BQCV), Israeli acute paralysis virus (IAPV), and chronic bee paralysis virus (CBPV)(2,3). In Mexico, the presence of DWV, IAPV and BQCV(4,5) has been reported in the high plateau region. Moreover, DWV and IAPV were identified in Varroa destructor mites, with DWV being the most prevalent in both, bee and mite samples(4). Not much is known about honey bee viral diseases in Mexico, and the information about them, is limited to a few regions of a few states. For Jalisco’s case, there are still no official reports about the distribution and levels of viruses in honey bee colonies of the state’s different beekeeping regions. It would be important to know this information because Jalisco is one of the foremost honey producing states in Mexico, ranking third in 2021 with 6,073 t(6).
For beekeeping purposes, Jalisco has been divided into six different regions that vary in topography and climate, including the regions of the Highlands, Center, North, Sierra Amula, South and Southeast. More than half of the state’s producers and hives are located in the South and Southeast regions(7). Because there is no information on the prevalence and infection intensity of honey bee viral diseases for the different regions of Jalisco, it seems relevant to conduct surveys to identify and quantify viruses in honey bee colonies from those regions, as well as to find out if there is a relationship between viruses and regions. Therefore, the objective of this study was to determine the prevalence and infection levels of the main viral diseases that affect adult honey bees, including DWV, BQCV, IAPV and CBPV, in samples of bees from colonies of six regions from Jalisco, Mexico.
Adult bee samples were collected during early spring, in the months of March, April and May 2018, from 12 to 16 colonies of each of the six beekeeping regions of Jalisco. In each apiary, three colonies were randomly selected and sampled. A total of 81 colonies from 27 apiaries were sampled, although data were obtained from only 79 colonies. Two samples of three bees each were collected from the entrance of each hive. The bees were introduced into 2 mL microfuge tubes containing RNAlater® (Thermo Scientific; Mississauga, ON, Canada) to preserve viral RNA. The samples were transported in coolers with freezing packs and were stored at -70° C until processed.
The molecular analyses to diagnose and quantify viral infections were conducted at the Honey Bee Research Centre, School of Environmental Sciences, University of Guelph, in Guelph, Ontario, Canada. First, the presence of DWV, BQCV, IAPV and CBPV, was determined by RT-PCR. RNA was extracted from three bees per sample with TRIzol (Fisher Scientific; Mississauga, ON, Canada), as per the manufacturer’s instructions. cDNA was synthesized with the RevertAidTM H Minus First Strand kit (Fermentas; Burlington, ON, Canada), following the manufacturer’s instructions.
The PCR reactions were carried out using a Master thermocycler (Eppendorf; Mississauga, ON, Canada). Each reaction contained 1.5 µL of 10x pH buffer for PCR (New England BioLabs; Pickering, ON, Canada), 1 µL of both primers (10 mM), 0.2 µL 5U/µL of Taq polymerase (New England BioLabs; Pickering, ON, Canada), 2 µL of cDNA and 8.8 µL of dH2O. The primer sequences and amplification cycles used were those described in previous studies for DWV(4,8), BQCV(9), IAPV(10) and CBPV(11). The PCR products were separated by electrophoresis on agarose gels and the amplified bands were photographed with a digital camera under UV light. Additionally, viral copies of DWV and BQCV were quantified with real time PCR (qRT-PCR). The other two viruses were not detected and therefore, not quantified. The calibration standard curve for DWV and BQCV was created using a 300 bp synthetic gene fragment or gBlock® (Integrated DNA technologies; Coralville, IO, USA) for each virus. The lyophilized of the synthetic genes (500 ng) were diluted with 50 µL of nuclease free dH2O to obtain an initial concentration of 10 ng/µL that was used for serial dilutions from 109 to 102 viral copy numbers.
The qRT-PCR reactions were done using a BioRad CFX96TM thermocycler (Bio-Rad Laboratories; Mississauga, ON, Canada) with PowerUp™ SYBRgreen™ (Supermix 2X) (Applied Biosystems; Foster City, CA, USA) on 96-well PCR plates (Hard-Shell®). The reactions had a final volume of 20 µL that contained the following. For DWV, 10 µL of Supermix 2X (Applied Biosystems; Foster City, CA, USA), 0.4 µL of both primers (200nM), 7.2 µL of nuclease free dH2O (Invitrogen; Burlington, ON, Canada) and 2 µL of cDNA or the synthetic gene dilutions. For BQCV, 10 µL of Supermix 2X, 0.8 µL of primers (400nM), 6.4 µL of nuclease free dH2O and 2 µL of cDNA or the synthetic gene dilutions. The primer sequences and amplification cycles were those described in previous studies for DWV(12) and BQCV(13).
The thermocycler software calculated the efficiency, determination coefficient (R2), and the slope of the viral RNA standard curve. To calculate the amount of viral RNA in the serial dilutions the following equation was used:
Number of viral RNA copies = (ng of synthetic gene) (6.022x1023) /(length of synthetic gene) (1x109) (650 D). Where: 650 D is the average weight of a base pair and 6.022x1023 is the Avogadro number(14). A graph using Ct values with the initial number of RNA viral copies and the number of DWV and BQCV copies of the samples was calculated using a regression equation.
To determine if there were differences between regions for viral prevalence, the data were analyzed with comparative tests for equality of proportions, using the Benjamini-Hochberg correction. Also, the data on infection intensity were subjected to Shapiro-Wilk and Bartlett tests to analyze the assumptions of normality and homoscedasticity, respectively. The data did not comply with the assumptions and thus, were log transformed and subjected to analyses of variance. When significance was detected, the regional means were compared with t tests using the Benjamini-Hochberg correction. All the statistical analyses were performed with the R 3.3.1 program (Foundation for Statistical Computing, Vienna, Austria).
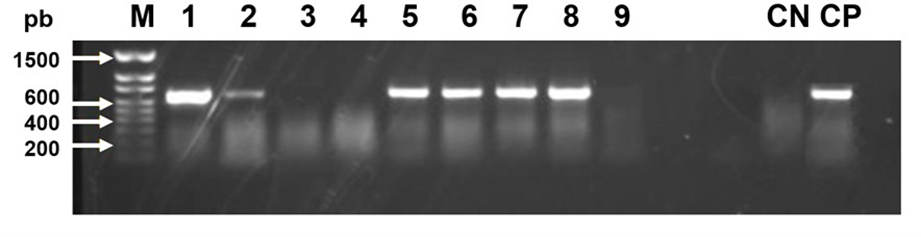
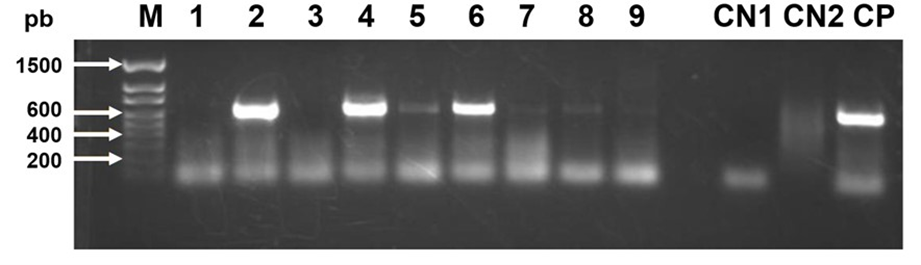
Two viruses were detected in the honey bee samples from all regions of Jalisco, BQCV (Figure 1) and DWV (Figure 2). The other two viruses, IAPV and CBPV, were not detected. Of the detected viruses, the prevalence of BQCV at the state level was 66 % and that of DWV was 38 %. The prevalence and intensity of the viral infections identified are shown in Table 1.
Figure 1: Photograph of an agarose gel that shows bands of 698 bp of black queen cell virus (BQCV) in columns 1, 2, 5, 6, 7 and 8. A honey bee gene (RpS5) is used as a control in the RT-PCR reaction
Figure 2: Photograph of an agarose gel that shows bands of 642 bp of deformed wing virus (DWV) in columns 2, 4 and 6. A honey bee gene (RpS5) is used as a control in the RT-PCR reaction
Table 1: Prevalence and mean intensity of viral infections that affect honey bee colonies in the state of Jalisco, Mexico
1 Number of viral copies per µg of RNA x 106
At the regional level, DWV prevalence in colonies from the Southeast and North regions was only 8 %, which was significantly lower than those of colonies from the South, Sierra Amula, and Center regions (P<0.05, Table 2). For DWV infection intensity, there were no significant differences between colonies of different regions (F5,73= 0.64, P=0.67).
Table 2: Prevalence and mean infection intensity of deformed wing virus (DWV) in adult workers of honey bee colonies in different regions of the state of Jalisco, Mexico
1 Number of viral copies per µg of RNA x 106.
ab Different literals indicate significant differences based on tests for equality of proportions using the Benjamini-Hochberg correction (P<0.05).
BQCV was detected in 42 to 81 % of the colonies from the different studied regions, but there was no significant difference between regions for BQCV prevalence (P>0.05, Table 3). However, for BQCV infection intensity, regions varied significantly (F5,73= 7.14, p< 0.01). For example, the South region had colonies with infection intensities significantly higher than those of colonies from the rest of the regions, except the Center region that was second for BQCV levels. The North region had the colonies with the lowest titers of BQCV infections.
Table 3: Prevalence and mean infection intensity of black queen cell virus (BQCV) in adult workers of honey bee colonies in different regions of the state of Jalisco, Mexico
1 Number of viral copies per µg of RNA x 106.
abcd Different literals indicate significant differences based on ANOVA and t tests using the Benjamini-Hochberg correction (P<0.05) of log transformed data.
In Mexico, there is little information about the presence of honey bee viral diseases, since viruses like DWV, IAPV, and BQCV were molecularly diagnosed for the first time just a decade ago in the Mexican high plateau(4,5), but nothing is known about the prevalence or infection intensity of these viruses in almost all states of the country. In northern Mexico, IAPV, DWV, sac brood bee virus (SBV), Kashmir bee virus (KBV), and filamentous virus (FV) were reported in colonies from the state of Chihuahua, but its prevalence and infection intensity were not determined(15,16). Therefore, the results of this study are a reference point for future research in Mexico’s regions of beekeeping importance.
In other countries of the Americas, several prevalence rates of honey bee viruses have been reported. However, most colonies of other countries have in common the prevalence of DWV and BQCV. For example, in Uruguay, 100% of the analyzed colonies were infected with DWV and BQCV(17,18). In Argentina and Chile, the most prevalent honey bee virus was DWV, which was detected in 35 y 37 % of the colonies sampled, respectively(19,20). In Cuba, DWV was the most prevalent virus, which was detected in 91 % of the colonies surveyed, but BQCV was not detected(21). In Colombia, both viruses were detected at a prevalence of 19.9 and 10.6 %, respectively(22). In North America, the prevalence of eight honey bee viruses was determined during six years in the USA, and in every single year, DWV was the most common of all viruses at a prevalence that ranged between 65 and 92 %, closely followed by BQCV with a prevalence range of 60 to 92 %(23).
Regarding viral infection intensities, with the exemption of reports from the USA and Canada, no study so far conducted in Central America, the Caribbean, or Mexico, has reported infection levels of honey bee viruses in different regions of a state, like this study does. Therefore, to the best of our knowledge, this is the first study to report infection levels of honey bee viruses at a regional level.
The prevalence of viruses varied between regions. DWV prevalence was significantly lower (8 %) in colonies of the Southeast and North regions than in other regions. Conversely, the prevalence of DWV in the regions South, Sierra Amula, and Center, was over 50 %. However, there were no differences for the intensity of infections of DWV between regions because viral infection levels were high in all regions. No differences in the prevalence of BQCV were found between colonies of different regions, however, their infection levels varied between colonies from one region to another. The colonies of the South region had higher BQCV infection levels than the colonies of the rest of the regions, except for the Center region. Regarding infection intensity, mean DWV infection levels were very high, with 4083.4 X 106 viral copies per µg of RNA, whereas for BQCV infection levels were relatively low, with 0.49 X 106 viral copies per µg of RNA. Thus, the intensity of DWV infection in honey bees from Jalisco was approximately 8,000 times higher than that of BQCV infection.
Some of the factors that could have influenced the differences in viral prevalence and infection intensity in honey bee colonies between Jalisco’s regions include environmental effects, bee genotype, and possibly different viral strains. Regarding climatic effects, it is known that DWV infections are more prevalent and intense in colonies located in temperate climates than in colonies established in tropical climates(24). The authors of the cited study proposed that this occurs because colder climates favor the transmission and replication of DWV and could reduce the immune responses of bees, making them more susceptible to the virus. The authors also found an effect of the interaction between climate and parasitism by V. destructor, a mite that is strongly related to the prevalence and infection intensity of DWV, since it not only serves as a vector of the virus, but the virus multiplies in the mite’s tissues(25,26). Therefore, colonies with greater V. destructor infestation rates tend to have higher DWV prevalence and infection intensity than colonies with low mite infestation levels(27). Furthermore, the genotype of bees varies with their degree of Africanization. It has been shown that the intensity of infection caused by DWV and BQCV is higher in colonies with bees of European mitotype or morphotype than in colonies with bees of African mitotype or morphotype(28). It is possible that the colonies that were less infected with viruses in this study, had a greater degree of Africanization than the more infected ones. However, this hypothesis would have to be investigated in future studies.
The high levels of DWV infection found in this study are concerning because if beekeepers neglect their V. destructor control measures, the prevalence and intensity of DWV infections could increase. It is known that together with Varroa parasitism, this virus can weaken colonies until they collapse(1). Therefore, it is essential to emphasize the importance of implementing an adequate control strategy for V. destructor infestations to keep DWV infections as low as possible in honey bee colonies. Regarding BQCV, although it had a high prevalence in colonies of most regions, its infection levels were low. However, this study was seasonal, and thus, studies would have to be carried out throughout an entire year and for several years, to confirm if it is a virus that could represent potential damage to the beekeeping industry in Jalisco.
In conclusion, the most prevalent honey bee virus in the state of Jalisco was BQCV, which was detected in 66 % of the colonies, while DWV was detected in 38 % of them. Infection levels of DWV were high (8,000 times higher than those of BQCV). The regions with the highest DWV prevalence were Center, South, Highlands, and Sierra Amula. Regarding the intensity of DWV infections, there were no significant differences between regions. There were also no significant differences between regions for BQCV prevalence, but there were for infection intensity of this virus. The regions with the highest infection levels were the South and Center regions. Additional studies are recommended with surveys conducted during different seasons of the year and for several years, to find out under what conditions and seasons, viruses could be harmful to the beekeeping industry, and to design control strategies.
Acknowledgments and conflict of interest
The authors thank the 42 beekeepers that kindly allowed the collection of honey bee samples from their colonies. Sara Dino, Ulises Nuño, Shaira Alvarado and Miriam Rangel, helped with sample collection. This study was partially financed by CUSur research funds granted to J.T. and by the University of Guelph Pinchin fund granted to E.G. The authors declare not to have conflict of interest.
Literature cited: