https://doi.org/10.22319/rmcp.v15i3.6541
Revisión bibliográfica
¿Resembrar o no resembrar? Factores que afectan el establecimiento de pastos de gramíneas en agostadero. Revisión
Aldo Torres Sales a
José Carlos Villalobos González b*
a Universidade Federal de Pernambuco. Pernambuco – Brasil.
b Texas Tech University. Davis College of Agricultural Sciences & Natural Resources, Goddard Building, Box 42125. Lubbock, TX 79409, United States.
* Autor de correspondencia: C.Villalobos@ttu.edu
Resumen:
Las plántulas tienen un papel importante en el mantenimiento de la diversidad, la productividad y los ciclos biogeoquímicos de los pastizales de cualquier zona geográfica. Sin embargo, poco se sabe sobre la influencia de los factores ambientales en el establecimiento de plántulas, así como sobre las diferencias en el desarrollo morfológico entre las especies. Comprender el establecimiento de plántulas se vuelve de crucial importancia para mejorar el éxito de la resiembra en pastizales. Esta revisión de literatura investigó los factores que causan fallas en el establecimiento de gramíneas en condiciones de pastizales. Germinar una semilla de pasto no es un gran problema si exixten condiciones ambientales óptimas. La clave es garantizar la supervivencia y crecimiento de estas plántulas hasta la madurez fisiológica. Los principales factores ambientales asociados con fallas en el establecimiento de plántulas son la humedad, temperatura ambiental, preparación de la cama de siembra, calidad de semillas, entre los principales son. Los estudios revisados mostraron que los pastos anuales tienen mayores tasas de crecimiento, sin embargo, presentan una menor asignación a las estructuras reproductivas en comparación con pastos perenes. Estas diferencias también promueven diferentes tasas de supervivencia de plántulas, con los pastos anuales presentando tasas más altas en comparación a pastos bianuales o perenes. Aparentemente, la causa principal de las fallas de las plántulas en el establecimiento está relacionada con la exploración de suelo de las raíces adventicias. Donde los informes describen que las plántulas emergen rápida y abundantemente en la mayoría de los pastos, pero las plántulas mueren entre las seis y diez semanas de edad. Popr lo que se deduce que la planta puede germinar y germinar las raíces primarias, sin embargo, por una razón desconocida, el pasto no brota la raíz adventicia.
Palabras clave: Pastizales, Raíz adventicia, Recuperación de pastizales, Resiembra de pastizales.
Recibido: 31/08/2023
Aceptado: 19/04/2024
Introducción
Los agostaderos son muy diversos, incluyen pastizales, sabanas, matorrales, desiertos, tundras, marismas y praderas. Los agostaderos cubren alrededor del 50 % de la superficie terrestre del mundo. Solo en América del Norte las estadísticas indican que 1,300 millones de hectáreas pueden clasificarse como agostadero(1). Durante el último siglo, el hombre ha incrementado las formas de utilizar los agostaderos debido a la expansión de las tierras de cultivo, la urbanización y, principalmente, la expansión de la producción ganadera. El uso excesivo de estos ecosistemas promueve un fenómeno no natural denominado “Degradación de los agostaderos”. Existen dos factores que pueden causar la degradación de los agostaderos: ciclos naturales liderados por sequías, y el segundo, relacionado con factores humanos que abordan el uso excesivo de recursos naturales.
Los efectos de las catástrofes climáticas y el uso de la tierra por el ser humano promueven el agotamiento de los nutrientes del suelo y la disminución de la retención de agua, lo que provoca una ruptura en la estructura del suelo y, por lo tanto, cambios en la composición de la vegetación, aumentando el porcentaje de plantas menos deseables para el ganado, como las plantas espinosas y de baja palatabilidad. Además, la intensificación del uso de la tierra sin una gestión adecuada de los agostaderos reduce la productividad de la biomasa, lo que puede conducir a una mayor expansión agrícola en zonas aún más marginales.
D’Odorico et al(2) mencionaron que, entre las actividades humanas que degradan los agostaderos, el sobrepastoreo es el principal factor de pérdida de diversidad de especies forrajeras en los agostaderos. El rápido aumento de la población ganadera mundial ha estado causando un uso excesivo de los agostaderos. El sobrepastoreo de los agostaderos reduce inicialmente la productividad de la biomasa, rompiendo todos los biociclos en los ecosistemas. En general, el sobrepastoreo convierte a los agostaderos en un lugar desértico (desertificación). Actualmente en todo el mundo, más de 680 millones de hectáreas de agostaderos se encuentran en alguna etapa de degradación, que es cinco veces más que el área de tierras de cultivo de Estados Unidos, o 100 veces el área de tierras agrícolas de México(3); Gaitán et al(4) afirman que el pastoreo continuo con cargas animales excesivas ha estado ocurriendo durante muchas décadas en todos los agostaderos de América.
La situación anterior ha alentado varios proyectos destinados a mitigar los efectos de la degradación de los agostaderos, incluida la resiembra de los agostaderos con especies de plantas nativas y exóticas. Es importante mencionar que actualmente, la mayoría de los proyectos recomiendan la resiembra de especies exóticas, creyendo en un aumento de la carga animal. No obstante, en algunas iniciativas, por ejemplo, en México y Argentina, la resiembra intenta restaurar el ecosistema a una condición cercana a la original, para así recuperar las condiciones del paisaje natural, el Plan Maestro de la Alianza Regional para la Conservación de los Pastizales del Desierto Chihuahuense en México y las Buenas Prácticas Ganaderas en Pastizales del gobierno argentino, se han vuelto obligatorios en los programas de resiembra el uso de solo especies nativas.
Johnson et al(5) afirman que la resiembra de gramíneas forrajeras nativas aumenta el porcentaje de especies de pastos deseables en el agostadero, aumentando el forraje disponible para el ganado y la vida silvestre y, en consecuencia, una mayor tasa de carga animal. Sin embargo, algunos proyectos de resiembra en varias partes del mundo (como México, Estados Unidos, Canadá y Argentina) están tratando de compensar la pérdida de diversidad vegetal utilizando especies de pastos nativos. No obstante, el alto costo y riesgo que implica el proceso de resiembra y la falta de conocimiento de los mecanismos fisiológicos en las plántulas parecen ser un gran obstáculo para el éxito de esta práctica. Además, Rector(6) mencionó que el alto riesgo de resembrar agostaderos está asociado con la imprevisibilidad temporal y espacial del patrón de lluvias en muchos agostaderos durante el periodo de establecimiento de las plántulas.
La probabilidad de éxito disminuye drásticamente en las zonas semiáridas debido a la corta temporada de lluvias. Rector(6) también afirma que la tasa de éxito en los programas de resiembra es más efectiva en zonas con precipitación anual superior a 800 mm/año. Por el contrario, en zonas con precipitación inferior a 350 mm anuales, la probabilidad de riesgo de fracaso en el establecimiento de las plántulas es superior al 80 %. De acuerdo con el Plan Maestro de la Alianza Regional(7), la resiembra de agostaderos promueve múltiples beneficios, tales como: 1) Aumento de la diversidad vegetal; mejorando la cantidad y calidad del forraje para el ganado; 2) Producción de semillas grandes o abundantes para la vida silvestre; 3) Promoción de una hermosa vista primaveral, y 4) Facilitación de la estabilización del suelo en áreas perturbadas. Si todas las condiciones ambientales (temperatura y humedad del suelo) se cumplen, se completará la germinación de una semilla de gramínea. El problema es asegurar la supervivencia y el crecimiento de estas plántulas hasta su establecimiento completo como plantas; en México la ocurrencia de sequías intraestacionales y heladas tempranas en los agostaderos afecta el establecimiento de las plántulas. Después de revisar un número considerable de trabajos, se consideró el fracaso del establecimiento de plántulas y el abordaje de las condiciones no ideales para germinar plántulas de gramíneas.
Orloff et al(8) señalan los tres principales factores asociados con fallas de las plántulas en los pastos de agostadero durante el establecimiento: condiciones ambientales inapropiadas, el tamaño de la semilla y factores genéticos. Sobre el factor genético, Esaú(9) y Tischler et al(10) reportan que el establecimiento de las plántulas se correlaciona con la capacidad de iniciar el crecimiento de las raíces adventicias y la posterior elongación de estas raíces en la etapa de plántula. En cuanto a los factores ambientales asociados con el establecimiento de las plántulas en gramíneas perennes, la mayoría de la literatura publicada mencionó cuatro factores como los más importantes para el establecimiento de las plántulas: 1) humedad inadecuada del suelo, especialmente en la superficie(11,12), 2) temperatura ambiental inadecuada, especialmente la temperatura del suelo(13), 3) competencia por la luz solar y los nutrientes entre las especies(14) y 4) profundidad de siembra(15). Respecto a los factores ambientales, Briske y Wilson(13) estudiaron la temperatura y humedad óptimas en plántulas de pasto blue grama y concluyeron que incluso con condiciones ambientales óptimas, algunos otros factores desconocidos también afectan el establecimiento de plántulas.
Aunque esta revisión tiene como objetivo discernir los desafíos de las plántulas durante el desarrollo estable, es esencial mencionar que también se deben superar algunas limitaciones de la semilla/germinación. El tamaño y peso de las semillas también se reportan como un factor que afecta el desarrollo de las plántulas(15). Maron et al(16) concluyeron que no solo el tamaño sino también el peso de las semillas influye en la supervivencia de las plántulas en las plantas de agostadero. Hyder et al(17) afirman que factores genéticos desconocidos también podrían promover fallas en el establecimiento de las plántulas. Además, otros trabajos(18) sugieren que los factores fotomorfogénicos asociados con la semilla y la corona podrían causar fallas. Varios enfoques han demostrado qué mecanismos afectan el establecimiento de las plántulas; sin embargo, todos concluyen que el desarrollo y extensión de las raíces adventicias es el proceso más importante asociado con el establecimiento de las plántulas. Por lo tanto, en esta revisión se discutieron los principales factores que causan fallas en el establecimiento de plántulas de pastos nativos en condiciones de agostadero.
Factores ambientales que afectan el establecimiento y la supervivencia de las plántulas
No existe un solo atributo ambiental que pueda explicar por completo el vigor de las plántulas en las gramíneas. Por lo tanto, las influencias ambientales en el establecimiento de plántulas en las gramíneas deben analizarse conjuntamente. La mayoría de la literatura indica que la interacción entre la humedad, la temperatura y la luz son los principales factores ambientales que afectan el éxito de la germinación de las semillas y la supervivencia de las plántulas.
Sluijs y Hyder(19) afirman que, en el pasto blue grama, la raíz adventicia crece fuera de las coronas de ahijamiento y se establece con éxito cuando el clima húmedo y nublado persiste durante 2 o 3 días después de la germinación. Pero si las raíces están expuestas a condiciones ambientales duras, las posibilidades de supervivencia son escasas(20). Es importante mencionar que cada sitio tiene una condición ambiental óptima para el establecimiento de las plántulas. El efecto de los factores ambientales en la germinación de las semillas de gramíneas de agostaderos ha sido ampliamente estudiado(21,22). No obstante, pocos estudios intentaron comprender el impacto del ambiente después de la germinación hasta el establecimiento completo (fase juvenil). En este segmento de esta revisión, se estará discutiendo el efecto de los factores ambientales en el establecimiento de las plántulas.
Temperatura
La temperatura parece tener más influencia en la germinación de semillas que en el establecimiento de plántulas. En general, la temperatura ambiental determina la tasa de desarrollo de todos los organismos. Snyman(23) afirma que la temperatura del suelo es el principal factor asociado con la emergencia de las plántulas porque controla la tasa de evaporación y transpiración en los ecosistemas. Se afirma que la temperatura ideal de crecimiento de las plántulas para las gramíneas de estación cálida está entre 25 y 30 °C(24). En contraste, para las gramíneas de estación fría, McGinnies(25) afirmó que una temperatura de alrededor de 20 °C promueve el mejor desarrollo radicular en estos grupos de especies. Mediante el estudio del efecto de la humedad y la temperatura en dos gramíneas y cuatro forbias nativas de pastizales de América del Norte, se concluyó que una temperatura inferior a 15 °C dio lugar a retrasos en el establecimiento de plántulas causados por una distribución ineficiente de las raíces en el suelo(26).
La temperatura del suelo parece ser más importante que la temperatura del aire. Hsu et al(24) sugirieron que la alta temperatura del suelo en los primeros 30 cm del suelo es un factor importante asociado con fracasos en la emergencia de las raíces. Por lo tanto, cuanto más rápido llegue la raíz a las capas más profundas del suelo, mayores serán las posibilidades de que la planta sobreviva y se establezca en el agostadero.
Hsu et al(24) afirman que la temperatura óptima del suelo para el crecimiento de las raíces para gramíneas de estación cálida oscila entre 9.4 y 11.4 °C. Sin embargo, esta temperatura suele ser más alta durante el verano en todos los agostaderos situados entre los paralelos 30º N y 30º S(27), lo que probablemente es uno de los muchos factores asociados con los fracasos en la resiembra en tierras secas. Briske y Wilson(13) mencionaron que, para un exitoso establecimiento de plántulas en tierras secas, las raíces de las plántulas de gramíneas deben desarrollarse rápidamente con el fin de evitar la exposición excesiva a altas temperaturas. La temperatura óptima para el brote y la raíz muestra ser diferente. La temperatura óptima para el crecimiento de los brotes para gramíneas de tierras secas es de alrededor de 5 °C inferior a la temperatura para un crecimiento óptimo de las raíces, lo que provoca un desequilibrio en cuanto a las condiciones ambientales(28).
Humedad
No hay duda de que la humedad tiene un papel esencial para las plantas en la etapa de plántula. La humedad parece no estar limitada en las tierras de pastoreo, que pueden recibir agua adicional mediante el riego. En cambio, en los agostaderos, la precipitación es la única fuente de agua. La humedad en el suelo no es un factor limitante para el crecimiento de las gramíneas en las tierras tropicales, ya que la humedad en el suelo de estas regiones está abundantemente disponible durante gran parte del año(29). No obstante, este es un factor limitante para el establecimiento de plántulas en ambientes semiáridos y áridos.
En cuanto al régimen de lluvias en Estados Unidos, Rajagopan y Lall(30) afirman que los agostaderos de los Estados Unidos al oeste de los 100º de longitud meridiana exhiben una considerable complejidad de distribución temporal y espacial, en comparación con los patrones de precipitación en la parte oriental del país. Por lo tanto, un análisis basado en la precipitación anual o mensual para las tierras secas no permite resultados concluyentes sobre la disponibilidad de agua para las plantas nativas.
Se mencionó que es necesario apreciar que la precipitación pluvial no implica que el agua de lluvia estará disponible al 100 % para las plantas(31). Por lo tanto, es esencial comprender los patrones hidrológicos y su implicación en el ecosistema. Así, este podría ser el paso inicial para comprender la influencia de los factores ambientales en el establecimiento de las plántulas. Algunos investigadores(32) abordaron la respuesta de las plantas con pulsos de precipitación. La teoría de los pulsos de precipitación sugiere que la frecuencia de las precipitaciones tiene la misma importancia que el volumen precipitado en alguna zona.
En un trabajo(33) se indica que es necesario tener dos días húmedos para que una semilla de pasto germine y cinco días húmedos para que la plántula se establezca en condiciones de agostadero. Pasado este tiempo, la plántula puede resistir hasta siete días secos consecutivamente. En tierras áridas, el crecimiento de las plantas se controla principalmente por la disponibilidad de agua en el suelo que por cualquier otro factor. El agua tiene una relación intrínseca con todos los aspectos del crecimiento de las gramíneas en las especies de agostadero, incluida la anatomía, la morfología, la fisiología y la bioquímica(34-36).
La clave para el establecimiento de plántulas en los agostaderos parece ser un equilibrio positivo en el agua disponible en el suelo. En otras palabras, debería haber más precipitaciones que evaporaciones en esa zona. Frasier et al(33) estudiaron el efecto de la sequía en las plántulas de sideoats grama y concluyeron que cinco días secos consecutivos promovieron la mortalidad de más del 50 % de las plántulas. La sequía y la desecación del suelo son los principales factores para los límites del establecimiento de plántulas en muchos ambientes(37). Se sugirió que existen diferencias en la supervivencia de plántulas entre especies de gramíneas durante la sequía. El desarrollo y la extensión de las raíces adventicias juegan un papel importante en el establecimiento de plantas y la exploración del suelo(38).
Al estudiar el pasto blue grama, se concluyó que la humedad óptima para el máximo desarrollo de la raíz adventicia es el 90 % de la saturación del suelo(13). Sin embargo, la raíz adventicia podría crecer lentamente en condiciones de bajo potencial hídrico del suelo. Harrington(39) estudió el efecto de la humedad del suelo sobre la supervivencia de las plántulas de arbustos en un pastizal semiárido en Australia y concluyó que para obtener éxito en el establecimiento de las plántulas fue necesario aplicar al menos 100 mm de riego suplementario durante el verano, tres veces después del riego previo a la siembra a principios de primavera y dos veces después del riego a finales de primavera. Es importante destacar que el autor alcanzó una supervivencia de plántula superior al 80 % en las parcelas de regadío en comparación con la supervivencia nula en las parcelas sin riego. Davis(40) afirma que la mayoría de las especies nativas del chaparral de California tienen plántulas adaptadas a la sequía que muestran raíces poco profundas y alta eficiencia de agua disponible. Se concluyó que las fallas en el establecimiento y desarrollo de las raíces de 12 especies de gramíneas de ocurrencia predominante en el chaparral de California ocurren durante las sequías en el verano(41).
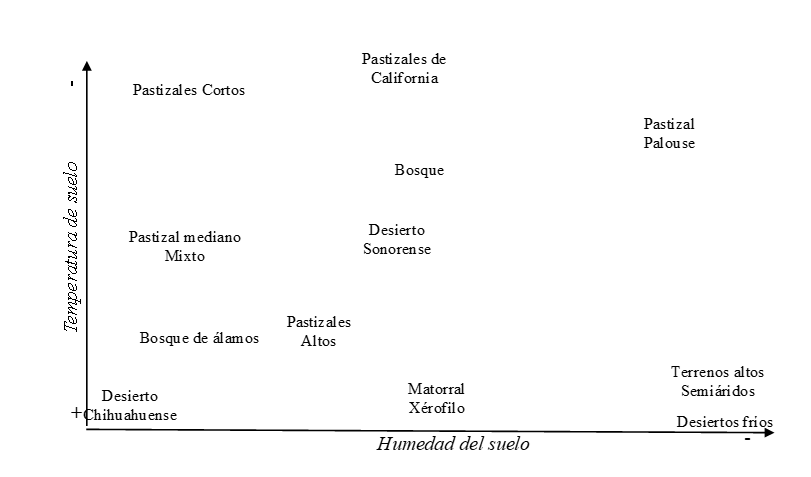
De acuerdo con algunos datos(1), la humedad y la temperatura en el suelo son los factores más importantes asociados con fracasos en el establecimiento de plántulas de gramíneas en agostaderos de América del Norte. Así, a partir de esta afirmación, se revisaron varios artículos que estudiaban la influencia de estos dos factores ambientales en los siete principales ecosistemas de agostadero de los Estados Unidos. La Figura 1 representa gráficamente el nivel de importancia de la humedad y la temperatura en el suelo reportado en la literatura científica asociada con fallas en el establecimiento de las plántulas de gramíneas en los agostaderos de América del Norte.
Figura 1: Importancia de la temperatura y la humedad del suelo asociadas con el riesgo de fallas en el establecimiento de plántulas de gramíneas en los pastizales de América del Norte *†
† La figura se basa en los resultados de 18 estudios en los 12 ecosistemas de pastizales de América del Norte.
*El nivel de importancia se ha determinado con base en una serie de informes científicos que asocian las fallas en el establecimiento de plántulas con la temperatura y humedad del suelo. Así, la cercanía de los ecosistemas (nombre) con la señal (+ y –) representa gráficamente el nivel de importancia de estas variables en el ecosistema.
En el desierto de Chihuahua, la combinación entre la humedad del suelo y la temperatura del suelo tiene un nivel similar de importancia en el establecimiento de plántulas de gramíneas de agostadero. Por otro lado, en los desiertos fríos y las tierras altas semiáridas se encontraron más reportes que demuestran una fuerte asociación entre los fracasos en el establecimiento de plántulas y la temperatura del suelo.
En los matorrales xéricos (p. ej., Matorral xerófilo y Mezquitales de México), la temperatura del suelo se reportó más que la humedad como causa de fracasos de plántulas en el establecimiento. Patrón similar al de las praderas de pasto alto y los parques de Aspen de Canadá. Para el resto del grupo, también se puede notar que, en las praderas de pasto alto y pasto medio, parece existir una combinación entre estos dos factores que aseguran un desarrollo exitoso de las plántulas de pastos de agostaderos.
Otros (luz, suelo y profundidad de siembra)
La luz también puede controlar el establecimiento de las plántulas porque una baja intensidad de luz reduce el tamaño de la hoja y la raíz. Pang et al(42) afirman que la sombra podía reducir la temperatura del suelo, pero no aumentó la supervivencia de las plántulas de pasto. Mediante el estudio del efecto de la sombra en la respuesta de crecimiento de cuatro gramíneas perennes del suroeste, se concluyó que las respuestas morfológicas, fisiológicas y de rendimiento fueron mayores en las plantas en condiciones de plena luz solar que en las plantas bajo diferentes niveles de sombra(43). Aunque la luz afectó el tamaño de las hojas y las raíces, esta variable no afectó el establecimiento de plántulas directamente.
En cuanto a las propiedades del suelo que afectan el establecimiento de las plántulas, la conductividad hidráulica parece ser la más importante, es decir, la capacidad de retención de humedad y la disponibilidad de agua para las plantas. Okami(44) declaró que la conductividad hidráulica es la variable del suelo más importante relacionada con el desarrollo de las plántulas de gramíneas, una vez que el 75 % del tiempo durante el establecimiento de la plántula, la plántula no depende del contenido de nutrientes en el suelo. Las características físicas del suelo, como la textura, la estructura, la densidad y la capilaridad, determinan la retención de agua en el suelo y la superficie de contacto entre la humedad del suelo y la semilla. Berti y Johnson(45) estudiaron el establecimiento de plántulas de pasto switchgrass en diferentes tipos de suelo, y concluyeron que la emergencia de las plántulas es un 25 % más rápida en suelos arenosos que en suelos arcillosos. Resultados similares indicaron que la textura del suelo afectó la emergencia de plántulas en alguna gramínea tropical(46).
La profundidad de siembra también se ha reportado como un factor que influye en el desarrollo de plántulas en gramíneas. Un informe(11) afirma que la profundidad de siembra afecta el establecimiento de plántulas de gramíneas de agostadero, especialmente durante la etapa de emergencia. Al analizar la influencia de la profundidad de siembra en la emergencia de plántulas de gramíneas nativas, se declaró que el pasto Bromegrass (Bromus inermis) sembrado a más de 1.3 cm de profundidad en un suelo franco arcilloso limoso disminuyó la emergencia y, en consecuencia, la supervivencia de las plántulas(47). Mientras que otros autores(11) estudiaron la influencia de la profundidad de siembra en la emergencia, la morfología y el establecimiento de los pastos big bluestem (Andropogon gerardii), Indiangrass (Sorghastrum nutans) y switchgrass (Panicum virgatum). Ellos concluyeron que los resultados obtenidos de estos experimentos no eran convincentes para afirmar que la profundidad de siembra afecta la supervivencia de las plántulas. En condiciones áridas, el pasto switchgrass tuvo la mayor emergencia cuando se sembró bajo riego previo a la siembra y siembra poco profunda(48). Anderson(49) afirma que las profundidades óptimas de siembra de pastos nativos del norte de Estados Unidos y sur de México están entre 6 a 12 mm, la diferencia dentro de este rango está en función de la especie de pasto y tipo de suelo.
Las diferencias en la tasa de utilización de las reservas de semillas pueden explicar por qué ciertas especies emergen a mayores profundidades de siembra que otras. Otros autores sugirieron que la profundidad óptima de siembra se correlaciona con la cantidad de hidratos de carbono que contiene la semilla(50-52).
Factores no ambientales que afectan el establecimiento de las plántulas
Las plantas tienen varias adaptaciones anatomorfológicas para competir más efectivamente entre sí por los recursos (luz, agua, nutrientes, suelo y aire). Harris(53) declaró que, en un concepto general, las gramíneas perennes tienen una ventaja competitiva natural sobre las gramíneas anuales, por lo que no es necesario que se siembren después de cada período de inactividad. Sin embargo, las plantas anuales tienen un establecimiento de plántulas rápidas.
Por lo tanto, al estudiar la influencia de la forma de crecimiento y la morfología de la planta en el establecimiento de las plántulas de gramíneas de tierras secas, los investigadores concluyeron que ninguna de las variable afectó la emergencia, la supervivencia y las tasas relativas de crecimiento en todas las formas de crecimiento estudiadas(54). Corroborando con los autores mencionados anteriormente, Larson et al(12) afirman que la variación en anatomía, morfología y fisiología entre semillas y plántulas explicó más del 90 % de la variación en la supervivencia acumulada de las gramíneas de agostadero, independientemente de las probabilidades de supervivencia de las plántulas o del patrón de precipitación.
Existen tres grandes factores no ambientales relacionados con el establecimiento de las plántulas: morfología, características fisiológicas y genéticas intrínsecas para cada especie (etapa serial) y competencia entre especies(21). A continuación, se presenta una breve revisión de cómo estos factores afectan el establecimiento de las plántulas en especies de gramíneas de agostadero.
Morfología
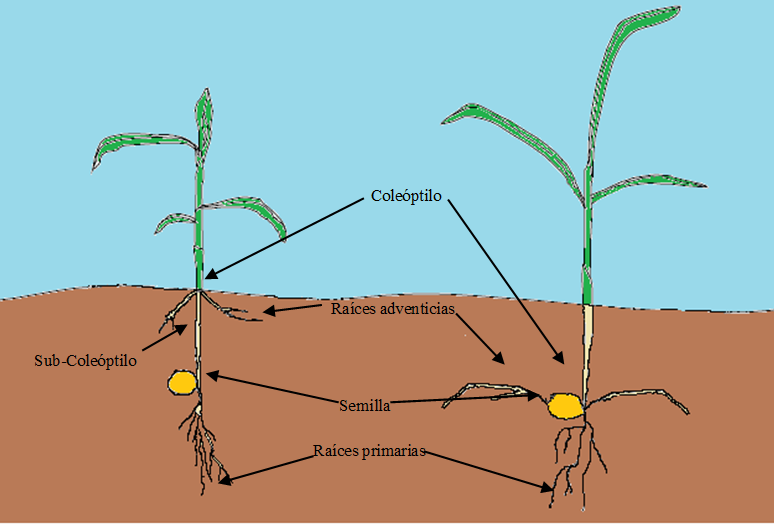
Las plántulas de gramíneas son hipogeas, lo que significa que el cotiledón en la mayoría de los casos permanece bajo tierra durante la germinación. La elongación del coleóptilo es diferente entre las gramíneas de estación fría y cálida. En las gramíneas de estación fría, el coleóptilo es largo con subcoleóptilo corto; por el contrario, las gramíneas de estación cálida tienen un coleóptilo corto y un subcoleóptilo corto (Figura 2).
Figura 2: Morfología de la plántula (Planta festudicoide derecha); izquierda (planta panicoide)
(Adaptado de Tischler et al 1989).
La extensión de los coleóptilos y subcoleóptilos tiene una importante papel en la emergencia de la plántulas porque cerca de la parte superior de estas estructuras se encuentran los puntos meristemáticos donde crecerá la primera hoja(55). Las raíces también son importantes en el establecimiento de las plántulas; se informó(56) que el sistema radicular de las plántulas de gramíneas consiste en raíces seminales y adventicias. Las raíces seminales comienzan a crecer inmediatamente después de la germinación; surgen directamente de una estructura en la semilla llamada nota escutelar. Las raíces seminales se dividen en raíces primarias y raíces laterales. Las raíces seminales son totalmente dependientes de niveles adecuados de contenido de agua en la planta joven, así como de la humedad en el suelo(57).
Después del desarrollo completo de las raíces seminales, la plántula comienza a liberar la raíz adventicia del nodo coleoptilar. Tischler y Voight(9) afirman que las raíces adventicias son consideradas el sistema radicular maduro. Algunos autores sugieren que el establecimiento de plántulas se asocia con el desarrollo de raíces adventicias(56).
Las raíces seminales comienzan a crecer después de 1 o 2 días húmedos consecutivos; una vez que las raíces seminales se desarrollan, las plantas comienzan a emitir la raíz adventicia. Esta fase se denomina etapa de transición. En otras palabras, la etapa de transición consiste en el desarrollo y la extensión de la raíz adventicia, y un debilitamiento y muerte de la raíz seminal(58). Hyder et al(17) afirman que en condiciones de campo en las que las plántulas de pasto blue grama fallan en la extensión de las raíces adventicias, las plántulas mueren entre las 6 a 10 semanas de edad. Se indicó que, para un establecimiento exitoso de plántulas, la tasa de elongación de las raíces adventicias debe ser lo suficientemente rápida como para mantener una parte de la raíz en el suelo húmedo antes de que se seque el suelo(13).
Newman y Moser(11) definen que el desarrollo de las raíces adventicias indudablemente controla el establecimiento de plántulas. Sin embargo, poco se sabe sobre cuántas raíces adventicias y la extensión de estas serán suficientes para afirmar cuando una planta se ha establecido. A pesar de que algunos autores asocian el establecimiento de plántulas con la etapa de transición, se mencionó el desarrollo y las actividades de las raíces de gramíneas cultivadas y se concluyó que las raíces seminales permanecen vivas y activas hasta el momento de la cosecha en especies de plantas de cultivo(57).
Después de examinar los informes sobre los sistemas radiculares de 14 gramíneas perennes, se concluye que en estas especies la raíz seminal creció profundamente y se extendió ampliamente, y permanecieron vivas y activas como órganos absorbentes durante cuatro meses de análisis experimental(13). La mayoría de los artículos revisados afirman que el desarrollo de las raíces adventicias en la etapa de transición determina el establecimiento de las plántulas, y la muerte de las raíces seminales es necesaria para el desarrollo de las raíces adventicias. No obstante, no existe un consenso sobre esta hipótesis, ya que algunos investigadores indican que las raíces seminales permanecen vivas durante mucho tiempo después de que la planta alcanza la fase madura.
Competencia entre especies
La competencia entre plantas se produce cuando las demandas de las plantas vecinas superan la oferta de recursos, induciendo el estrés y entonces la muerte de las plantas. La finalización de la planta podría ocurrir en dos niveles, entre individuos de la misma especie (interespecie) o entre individuos de diferentes especies (intraespecie). Sea cual sea el nivel, la competencia afecta la disponibilidad de recursos ambientales para las plantas. Se reportó(59) que las densidades iniciales y el momento del establecimiento promueven cambios en la dinámica de la competencia de las plantas, ya que conducen a asimetrías en el tamaño de las plantas y captura de recursos. Las plantas de agostadero tienen muchas adaptaciones (morfológicas, anatómicas, fisiológicas y fenológicas) que las adaptan a un lugar en el ecosistema. Por lo tanto, la comprensión del efecto de la competencia entre las especies es un requisito básico para aumentar las posibilidades de éxito de la resiembra de agostaderos.
Los rodales de gramíneas perennes tienen una ventaja competitiva natural sobre las gramíneas anuales. Ya que no es necesario que comiencen desde la semilla después de cada período de inactividad(24). Ries y Svejcar(60) informaron que las plántulas de plantas anuales invaden fácilmente y se establecen en sitios perturbados. La fenología radicular favorable es una de las estrategias adaptativas que permite esta capacidad competitiva superior.
Algunos autores(61) definen cinco características fenológicas más importantes de las plántulas con respecto a relaciones competitivas en plantas jóvenes. Las características son: (1) fácil germinación, (2) crecimiento inicial precoz de las raíces, (3) extensión rápida de la raíz-contacto con el suelo, (4) fácil interrupción de la latencia, y (5) la supervivencia de la sequía.
La competencia de las plantas también podría dividirse de acuerdo con la zona donde se produce. En esta clasificación, podemos dividir la competencia en dos niveles: por encima y bajo tierra. Varios estudios han proporcionado evidencia de que en ambientes áridos la competencia en el subsuelo es más importante que en la superficie(62).
Harris y Wilson(61) sugirieron que en áreas donde la estación de humedad favorable coincide con la estación de bajas temperaturas, la capacidad de las plántulas para continuar el crecimiento radicular a bajas temperaturas puede ser un factor decisivo en el resultado de la competencia entre especies. También estudiaron el efecto de la humedad del suelo durante el establecimiento de plántulas de gramíneas forrajeras de estación fría a bajas temperaturas, concluyendo la existencia de diferencias en el crecimiento de las raíces adventicias, donde el Bromus tectorum y Taeniatherum asperwerum tuvieron más éxito que las plántulas de Agropyron spicatum.
Las especies invasoras han mostrado ser más eficientes en el establecimiento de plántulas que las especies nativas. Se reportó(63) que las especies invasoras germinan más rápido que las especies nativas, obteniendo una ventaja en la competencia por la luz, espacio y la humedad. Como se mencionó, se necesitan más estudios para saber cómo se produce la interacción entre plantas en la etapa de plántula en gramíneas nativas. Es importante mencionar que el primer paso en la recuperación de tierras es eliminar el banco de semillas.
Etapa serial
Varios estudios han demostrado innegablemente que las plantas anuales tienen una mayor tasa de crecimiento de plántulas(55,64) y una mayor asignación a las estructuras reproductivas(65,66) en comparación con las plantas seriales medias y de sucesión tardía. Newman y Moser(11) compararon el desarrollo de plántulas entre 12 especies de gramíneas. Ellos concluyen que las plantas anuales emitieron la primera hoja más rápido que las especies seriales tardías. Las especies seriales tempranas parecen asignar más energía al desarrollo de los brotes que al sistema radicular. Por el contrario, las especies seriales medias y tardías parecen asignar más energía al desarrollo de las raíces. Ellos concluyen que, si la proporción de brotes es mayor que la de raíces en la plántula, mayores son los riesgos de fracaso en el establecimiento causado por la acidificación de las plántulas o un control ineficiente en la absorción de agua(56).
Importancia del patrón de raíces seminales y adventicias en el establecimiento de la planta (implicaciones en la supervivencia de las plántulas)
Como se mencionó, el establecimiento y la supervivencia de una planta están intrínsecamente correlacionados con la aparición y extensión de las raíces seminales y adventicias(56). Otro trabajo(67) apoya la idea de que una planta no puede considerarse como establecida hasta mostrar un desarrollo plausivo de raíces adventicias, lo que permitirá extenderse hacia abajo, atrapando la humedad en los niveles profundos del suelo. Para facilitar la comprensión de la secuencia cronológica de los eventos morfológicos que ocurren en una planta hasta que el desarrollo de la raíz adventicia, se utilizó la descripción mencionada por Whalley et al(68). El autor divide el crecimiento de las plántulas en tres etapas: la etapa heterótrofa, una etapa transitoria, y la etapa autótrofa.
La etapa heterótrofa comienza cuando la semilla entra en contacto con el agua estando en esta etapa independientemente de otros factores ambientales, lo que significa que la planta utiliza en su metabolismo las reservas energéticas (almidones) almacenadas en la semilla. Después de esta fase, la planta todavía no tiene un tejido fotosintético. Hyder et al(17) afirman que las plantas superan fácilmente esta etapa en condiciones de campo. La mayoría de los investigadores consideran erróneamente el porcentaje de semillas germinadas como un indicador del establecimiento de plántulas. Hyder et al(17) afirman que menos de ocho por ciento de las semillas germinadas alcanzarán la fase adulta. La siguiente etapa transitoria, como el nombre sugiere, es una evolución de transición a una fase fotosintética. En esta etapa, las plantas comienzan la fotosíntesis, pero aún utilizan las reservas de energía de las semillas para la expansión de los sistemas radiculares y la formación de nuevas hojas. Esta fase se puede dividir en tres subetapas con base en el desarrollo del sistema radicular.
En la primera fase, la plántula solo muestra raíces seminales. El sistema de raíces seminales consta de una a cinco raíces que se desarrollaron a partir de la radícula y dos pares de raíces laterales. Algunos investigadores(21) afirman que las raíces seminales solo son capaces de absorber agua; mientras que todos los nutrientes necesarios para la plántula provienen de las reservas de las semillas. El grueso de las raíces seminales tiene una capacidad limitada para absorber y translocar agua en la planta. En la segunda fase de esta etapa, las plantas comienzan a emitir las raíces adventicias de los nodos de la corona. Algunos autores reportan este tipo de raíces como raíces verdaderas porque pueden absorber la humedad y los nutrientes para la planta. En esta fase, las plántulas muestran estos dos tipos de raíces. Poco se sabe sobre la interacción entre estos tipos de raíces. La tercera etapa a menudo se marca por el debilitamiento y la muerte de las raíces seminales y el fortalecimiento, extensión y consolidación de las raíces adventicias como raíces verdaderas. Haling et al(69) afirman que las raíces seminales persisten solo un corto tiempo después de la germinación, y su lugar es ocupado por las raíces adventicias. Algunos autores afirman que la tercera etapa de la plántula es el momento más importante en el establecimiento de plántulas(40,70). Si la planta comienza a perder sus raíces seminales antes de que las raíces adventicias alcancen una penetración profunda razonable en el perfil del suelo, hay una alta probabilidad de fracaso en el establecimiento.
Debido a una mala comprensión de las bases fisiológicas de las plántulas, la mayoría de los administradores de agostaderos asocian el establecimiento de plántulas con el desarrollo de brotes. En gramíneas nativas americanas se concluyó que la etapa de desarrollo radicular no coincidió con la etapa de desarrollo de brotes entre especies, lo que significa que solo un simple análisis visual no puede ser considerado un buen indicador de éxito en la emergencia de plántulas de gramíneas americanas(11). Así, se afirmó(9) que, para evaluar el éxito del establecimiento de plántulas, también son importantes variables como el tamaño y la edad en la plántula de pasto Klein y deben medirse adicionalmente al análisis visual.
En cultivares seleccionados del pasto big bluestem, con base en el peso de los brotes y numero de macollos, concluyen que el número y el peso de los macollos de las plántulas no son buenos indicadores del éxito del establecimiento de las plántulas(71). El desarrollo y la extensión de las raíces adventicias también parecen estar asociados con factores genéticos. Se reportó(72) que la variabilidad genética pareció correlacionarse con la elongación de raíces adventicias en el pasto blue grama, y que, en plantas bajo las mismas condiciones edafoclimáticas, mostraron una amplia diversidad de desarrollo que puede explicarse por factores ambientales.
Por el contrario, Chen et al(57) después de investigar los sistemas radiculares de varios cultivos, concluyeron que la raíz seminal permanece viva y activa hasta el momento de la cosecha, ésta puede ser la razón del éxito en el establecimiento de la mayoría de los cultivos cosmopolitas. Sin embargo, algunas especies de gramíneas parecen moverse en la misma dirección. Sánchez-Valdés et al(73) afirman que la raíz seminal de una planta de ryegrass permanece funcional durante toda la vida del planta. Mismos resultados se informaron para otras especies(74).
Literatura citada: