https://doi.org/10.22319/rmcp.v15i2.6539
Revisión
Contribución de gramíneas forrajeras a la fijación biológica de nitrógeno y su respuesta a la inoculación de diazótrofas. Revisión
Dania Fonseca López a
Nelson Vivas Quila b
Raúl Cuervo Mulet c
Carlos Eduardo Rodríguez Molano d*
a Universidad Santo Tomás. Centro de Investigación en Ciencia y Tecnología para el Desarrollo Sostenible. Tunja, Colombia.
b Universidad del Cauca. Facultad de Ciencias Agrarias. Grupo de Investigación NUTRIFACA. Popayán, Colombia.
c Universidad de San Buenaventura. Facultad de Ingeniería. Cali, Colombia.
d Universidad Pedagógica y Tecnológica de Colombia UPTC. Facultad de Ciencias Agropecuarias. Tunja, Colombia.
* Autor de correspondencia: carlos.rodriguez@uptc.edu.co
Resumen:
El uso de insumos de origen químico ha generado pérdida de la diversidad microbiana que interviene en el ciclo del N como bacterias diazótrofas, que son inhibidas por saturación de los receptores encargados de activar la nitrogenasa. La fijación biológica de nitrógeno (FBN) en gramíneas forrajeras puede ser utilizada como servicio ecosistémico. El objetivo de este artículo de revisión fue analizar la contribución de gramíneas forrajeras a la FBN y su respuesta a la inoculación de diazótrofas no simbióticas para encontrar oportunidades de estudio. El análisis de la información se realizó a partir de la metodología prisma de revisiones sistemáticas y meta-análisis. Se destaca que las principales especies forrajeras que contribuyen con la FBN corresponden a Brachiaria sp. y Pennisetum sp. La inoculación de Azospirillum sp. ha generado efecto promotor de crecimiento en gramíneas, pero, la respuesta del forraje inoculado depende principalmente de la sinergia entre planta – bacteria presentándose efectos neutros, antagónicos y positivos.
Palabras clave: Fertilización, Fijación nitrógeno, Forraje, Nitrogenasa, Pastos.
Recibido: 23/08/2023
Aceptado: 21/12/2023
Introducción
En los sistemas ganaderos la alimentación de los animales resulta viable económicamente cuando la ración está conformada principalmente por forraje. Pero, es necesario producir pasto en un escenario ecoeficiente para compensar la huella ambiental ocasionada por la ganadería, teniendo en cuenta que en Colombia ocupa el 80 % del suelo con vocación agrícola(1). Las estrategias propuestas incluyen el uso de especies forrajeras mejoradas, la diversificación del sistema(1) y el aprovechamiento de fenómenos naturales como la fijación biológica de nitrógeno (FBN)(2). Este, es un proceso en que diazótrofas transforman nitrógeno (N) atmosférico en amonio a partir del complejo enzimático nitrogenasa, y aporta cerca del 62 % lo que equivale a 11.29 millones de toneladas (Mt) de nitrógeno al año que ingresa al ecosistema agrícola Latinoamericano, mientras que la fertilización química contribuye aproximadamente con 6.81 Mt N al año(3). La FBN es un recurso que puede ser utilizado como herramienta tecnológica para reducir la aplicación de fertilizantes nitrogenados de origen sintético que tienen baja eficiencia (aproximadamente 40-50 %), y contribuyen con la emisión de gases efecto invernadero (amonio, amoniaco y el óxido nitroso)(4) y salinización del suelo(5). Sin embargo, poco se conoce sobre el aporte de las gramíneas forrajeras a la FBN y las especies bacterianas con mejor efecto productivo. Por esto, el objetivo de este trabajo fue analizar la contribución de gramíneas forrajeras a la FBN, y su respuesta a la aplicación de biofertilizantes constituidos por bacterias diazótrofas no simbióticas puras, y en consorcio a partir de una revisión sistemática de literatura para encontrar oportunidades de estudio.
Se utilizó la metodología prisma de revisiones sistemáticas(6), las bases de datos consultadas fueron Scopus y Web of Science; para la búsqueda de información se establecieron los siguientes criterios: a) especificidad, a partir del uso de operadores boléanos, b) sensibilidad, con descriptores CAB; c) exhaustividad, a través de la comprobación de los descriptores de interés. La estrategia de búsqueda fue a partir de las rutas: TITLE-ABS-KEY (“Biofertilizer”) y TITLE-ABS-KEY (“Biofertilizer and Grass”). Con la búsqueda general se encontraron en total 6,813 registros entre las bases Scopus (n=4,621) y Web of Science (n= 2,192). La búsqueda se limitó a los conectores boléanos “Biofertilizer and Grass” a partir de los cuales se encontraron 128 registros (Scopus: 84 registros y Web of Science: 44 registros) que fueron importados al software Mendeley y se agruparon por años; el análisis se limitó al periodo 2012-2022 (n= 80 registros), luego se eliminaron los documentos duplicados (n= 2 registros). Se incluyeron en el análisis artículos donde se evaluó el efecto de la aplicación de biofertilizantes en forrajes o la contribución de nitrógeno fijado por estas plantas. Se excluyeron publicaciones con un título por fuera de la búsqueda de interés (n= 5) y con información únicamente descriptiva que no cumplían los criterios de inclusión (n= 13 registros). Cada registro se revisó de forma independiente por todos los autores para un total de 50 estudios incluidos dentro de la revisión. Se definieron como resultados del análisis: a) Nitrógeno fijado por gramíneas forrajeas, b) Biofertilizantes aplicados y su efecto en gramíneas forrajeras. Los datos de interés de estudio (Nitrógeno fijado y efecto en planta) fueron tabulados y agrupados por tema para medir su efecto. Se realizó un análisis de regresión no lineal con el número de los registros obtenidos a partir de los modelos sigmoidal 3,4, Gompertz 3, y Hill 3. Los modelos con mayor ajuste se seleccionaron de acuerdo con el valor de significancia y ajuste del coeficiente de determinación para establecer la tendencia global del área de interés.
Fijación biológica de nitrógeno en gramíneas forrajeras
En esta revisión se identificó que la prueba de elección para determinar el nitrógeno fijado por gramíneas forrajeras es abundancia natural de 15N(7). En las principales investigaciones donde se reporta N fijado por forraje, se destaca que la tasa de fijación de N difiere entre especies (Cuadro 1). Esto tiene una relación directa con las poblaciones de bacterias diazótrofas que interactúan con cada tipo de forraje, en Brachiaria sp. se estima aproximadamente de 102 a 108 UFC g−1 suelo(8). Mientras que en Pennisetum sp. se reporta que la población bacteriana diazótrofa es de 102 a 106 UFC g−1 suelo(9).
Cuadro 1: Algunos reportes de especies forrajeras que contribuyen con la fijación biológica de nitrógeno según análisis de revisión
Fuente: elaborado a partir de citas indicadas.
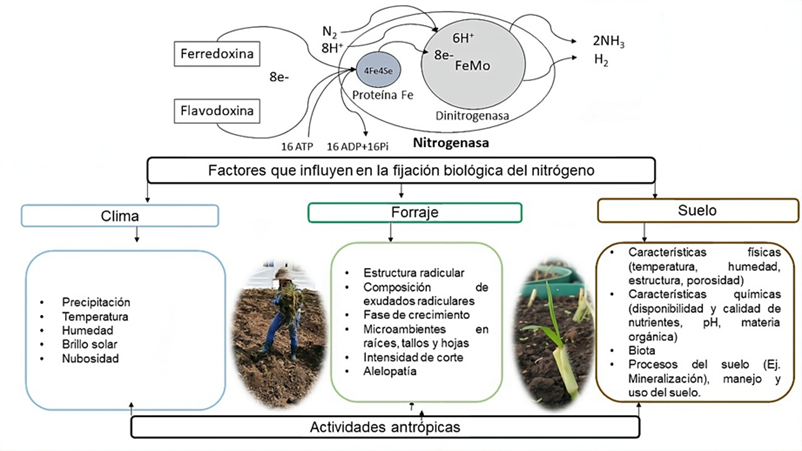
Se encontró que los principales géneros bacterianos que persisten en rizosfera y tejido vegetal de Brachiaria sp., Pennisetumm sp., Megathyrsus sp. y Panicum sp., corresponden a Enterobacter sp. (6 %)(10), Azospirillum sp. (25 %)(12,13,14), Azotobacter sp., Bacillus sp. (14 %)(2,15), Herbaspirillum sp. (11 %), Burkholderia sp. (8 %)(14), Bradyrhizobium sp. (6 %), Klebsiella sp. (5 %)(11,16), Sphingomonas sp. (4 %)(17), otras (2 %). Aunque, su distribución en raíces, hojas y tallos varía por especie forrajera, localidad y tipo de suelo(18). Estos microrganismos no causan modificaciones estructurales en la planta y están codificados con el gen nifH(4). El proceso de FBN lo realizan en sitios con menor saturación de oxígeno para evitar la inactivación de la nitrogenasa, como en las arcillas o a través de la reducción en la concentración de oxígeno intracelular a partir del incremento en la respiración celular(19). Durante la reacción, ocho electrones son bombeados a alta velocidad desde un agente donante (ferredoxina o flavodoxina) hacia el complejo enzimático nitrogenasa constituido por las metaloenzimas dinitrogenasa reductasa o proteína Fe codificada por el gen nifH y la metaloenzima dinitrogenasa codificada por los genes nifD y nifK(20). La dinitrogenasa reductasa trasfiere cada electrón a la dinitrogenasa y son almacenados en el cofactor FeMo, sitio de unión del N hasta que es reducido a NH3, de este modo, se consumen 16 ATP y se producen 2 mol de amonio y 1 mol de H2 por cada molécula de N fijada(21). Como resultado de la revisión, se encontró que las diferencias en los rangos de nitrógeno fijado entre forrajes y especies del mismo género está determinada principalmente por los factores: planta, suelo, actividades antrópicas y clima (Figura 1).
Figura 1: Factores que influyen en la fijación biológica del N en gramíneas forrajeras
Fuente: elaborada a partir de las citas(2,5,11,18,21,22).
Efecto del clima en la FBN de forrajes
Aunque son escasos los estudios en los que se analiza el efecto del clima sobre el proceso de FBN, se destaca que la nubosidad influye negativamente en este proceso, por la menor disponibilidad de fotoasimilados que se producen en las hojas y se distribuyen hacia las raíces para la formación de rizoexudados(23). La mayor producción de fotoasimilados parece tener una relación directa con la persistencia de diazótrofos inoculados, lo que favorece su efecto; por ejemplo, con la aplicación de Azospirillum brasilense en Urochloa brizantha se ha observado que al comienzo de la estación seca en la cual incrementa la radiación solar, la masa de las raíces de plantas inoculadas fue un 27 % mayor que en plantas no inoculadas, y aunque durante la época de transición disminuyó la producción de pasto, en plantas inoculadas se redujo en solo 7 % y su altura incrementó un 16 % respecto a plantas no inoculadas, por la mayor absorción de nutrientes(12). Respuestas similares se reportan con la aplicación de Bacillus sp. en Megathyrsus maximus(24).
Garantizar la persistencia de comunidades diazótrofas puede reducir la dependencia de fertilización nitrogenada(13); sin embargo, en época lluviosa la fijación de N por efecto bacteriano puede disminuir quizás por el arrastre de los microorganismos(1, 9). Esto puede explicar por qué se reporta que al final de la estación húmeda la biomasa radicular disminuye en 15 % en plantas inoculadas y se presenta menor contenido de N en las hojas respecto a plantas fertilizadas con N(12).
En ambientes deficientes de N la FBN incrementa como una respuesta de control cuando hay bajas tasas de mineralización(4,25). De este modo se reporta mayor N acumulado en otoño que en primavera por efecto de una menor temperatura en los forrajes Axonopus affinis (37.6 kg N ha-1.), Paspalum notatum (27.7 kg N ha-1.) y Andropogon lateralis (1.6 kg N ha-1) estimándose que en promedio el porcentaje de N proveniente de la FBN es de 33 %, 22 % y 25 % respectivamente(2).
Efecto del suelo en la FBN de forrajes
Las características del suelo también influyen en la FBN(22) destacan mayor diversidad de poblaciones diazótrofas en suelos con alta materia orgánica. La persistencia de estos microorganismos está modulada por el tipo y calidad de nutrientes en el suelo(22) explican, que los diazótrofos incrementan su actividad con presencia de hierro (Fe), molibdeno (Mo) y vanadio (V) debido a que estos elementos pueden intercambiarse para hacer parte de la estructura de la nitrogenasa. Esta enzima, al inactivarse con el oxígeno, requiere de micrositios anaerobios para catabolizar la fijación de nitrógeno, por esto, parece que en suelos arcillosos hay mayor movilización química, mineral, y eventualmente mayor FBN(21).
Efecto de las actividades antrópicas en la FBN de forrajes
El suelo es un sistema que se autorregula naturalmente, pero, cambios bruscos en sus características por actividades antrópicas de manejo (labranza, fertilización) y uso (pastos permanentes con y sin intervención, ganadería), causan desbalance en las comunidades bacterianas, ya que alteran la estructura de los poros, la disponibilidad de elementos, el contenido de carbono orgánico y el pH, factores que determinan la riqueza, uniformidad y diversidad de microorganismos(2,22).
La aplicación excesiva de Ca, nitrato y N durante la fertilización tiene un efecto negativo en las poblaciones diazótrofas(17). La principal causa se relaciona con el pH del suelo(12,22); variaciones en 1.5 del valor del pH del suelo puede reducir el crecimiento de los microorganismos hasta en 50 % en suelos con un pH entre 5 y 7(12,22). Se reporta, la inhibición en el crecimiento de algunas poblaciones microbianas, como Azotobacter, Azospirillum, Herbaspirillum y Gluconoacetobacter diazotrophicus con altas dosis de fertilización con N(2,10,26), por ejemplo, con la aplicación de 430 kg N ha-1 en B.brizanta y B. ruziziensis(27). Sin embargo, el tipo y cantidad de fertilizante aplicado influye en la abundancia y diversidad de las poblaciones microbianas; se ha observado incremento de metanótrofos con aportes mayores de 200 μg N g-1 de amoniaco al superarse el sitio activo del amonio monooxigenasa(28). En general, la modificación estructural de la comunidad bacteriana es un mecanismo natural de control del estado de nitrógeno en el suelo(2).
Efecto del factor planta en la FBN de forrajes
Las características morfo fisiológicas de las gramíneas generan microambientes disímiles en hojas, tallos y raíces, que promueven el crecimiento selectivo de miembros de la población bacteriana durante la fase de crecimiento(4). En la fase temprana, es mayor la actividad de poblaciones diazótrofas rizosféricas por incremento en las rizodeposiciones como mecanismo de recuperación de la planta posterior al pastoreo(13). La interacción entre las bacterias diazótrofas y la planta, sucede a partir de las rizodeposiciones que incluyen varias moléculas como azúcares, polisacáridos, ácidos orgánicos inorgánicos, aminoácidos, vitaminas, flavonoides, sideróforos, péptidos, proteínas y ácidos grasos(29). Estas señales químicas controlan las interacciones que suceden en el suelo y son las responsables de promover el crecimiento selectivo de miembros de la comunidad rizosférica y permiten el desplazamiento de las bacterias hacia la raíz de la planta y los pelos radiculares(24). La diversa capacidad funcional de las bacterias diazótrofas permite modular la respuesta de crecimiento del forraje y generar interacciones positivas, negativas o neutras. A continuación, se discuten los principales hallazgos encontrados en relación con la respuesta de forraje con la inoculación de diazótrofos.
Biofertilizantes constituidos por diazótrofas que han sido utilizados en gramíneas
A partir de 1985 se reportan los primeros estudios científicos en el área de biofertilizantes aplicados a forrajes, aunque históricamente es una práctica que data del año 500 A.C. originaria de la India, país que continúa liderando los avances científicos con un 30 % de participación mundial seguido de Brasil (10 %) y China (8.8 %). En el área de biofertilizantes aplicados en forrajes se destacan autores como Gupta et al(4), Li H et al(15) y De Sousa et al(30). Se estima un rápido crecimiento en el área con un punto de inflexión para el año 2034 (Cuadro 2), proyección que muestra la existencia de oportunidades de estudio que van ligadas con el fenómeno de cambio climático y el reto de utilización de estrategias de fertilización sostenibles que reduzcan la aplicación de productos químicos obtenidos por la quema de combustibles fósiles como la urea.
Cuadro 2: Modelos de regresión no lineal obtenidos para las búsquedas “Biofertilizer” y “Biofertilizer and grass”
Fuente: Elaboración propia.
La tendencia de uso de biofertilizantes en forrajes es sigmoidal con un punto de inflexión hacia el año 2029 como se observa en el modelo logístico con mayor ajuste que obtuvo un valor Durbin Watson cercano a 2(7), aunque, la predicción por los modelos Gompertz y Hill es anterior, pero con un menor ajuste (R2), por lo tanto, no predicen un comportamiento confiable (Cuadro 2). El punto de inflexión se asocia con la fase de rápido crecimiento de la tecnología y corresponde al valor máximo de la curva a partir del cual se prevé empiezan a disminuir las publicaciones relacionadas con biofertilizantes. Estas predicciones con alta variación se relacionan con áreas de aplicación en creciente desarrollo, y es que la fertilización orgánica empieza a cobrar importancia en el sector ganadero por el alza en el costo de los fertilizantes químicos.
A partir del análisis de revisión se encontró, que los biofertilizantes utilizados en pastos han sido aplicados por inoculación de semilla en el producto durante 30 min a 24 h seguido de un tiempo de secado previo a la siembra(2,31) o por aspersión en dosificaciones que varían entre 200 – 500 ml de inoculante ha-1 diluido en agua al 0.1 - 1.3 % en una concentración mínima de 106 UFC ml-1 o 106 UFC g-1(32-36).
La inoculación de microrganismos puede modificar el desarrollo del forraje con alta variabilidad entre géneros y cepas aplicadas o incluso no causar efecto, o generar una respuesta negativa(35) (Cuadro 3). Cuando los biofertilizantes se han aplicado junto con una fuente de N sintético, se han logrado respuestas superiores o equivalentes a la aplicación del 100 % del requerimiento de N por una absorción más eficiente, reduciéndose las pérdidas de N causadas por lixiviación hasta en un 95 %(36). Los mejores resultados en términos productivos y económicos se han observado con la aplicación combinada del inoculante y N(36-40).
Cuadro 3: Algunos estudios del efecto de la aplicación de diazótrofos en gramíneas forrajeras
Fuente: elaborada a partir de las citas indicadas.
La respuesta positiva de la planta con la inoculación de diazótrofos se da por dos condiciones principalmente, primero por favorecer la disponibilidad de nitrógeno en el suelo que es un elemento que hace parte de las proteínas, aminoácidos, ADN, ARN, citocromos, ácidos nucleicos y la clorofila(2,21); y segundo por la producción de metabolitos secundarios de origen bacteriano como: a) auxinas que están involucradas en el crecimiento, diferenciación y división celular(16), b) giberelinas, que son hormonas que intervienen la regulación de la división y el alargamiento celular, en la germinación de semillas, aparición de yemas y en el crecimiento de los tallos(48), c) citocinas que están relacionadas con la regulación del crecimiento celular(48), d) sideróforos que son compuestos que pueden unirse al hierro dejándolo disponible para ser utilizado en procesos metabólicos(26) y e) biosurfactantes que son agentes químicos que forman micelas y permiten mejor interacción entre la membrana de los microorganismos y los nutrientes disueltos en el suelo y en las rizodeposiciones(49).
De estas biomoléculas las más estudiadas son las auxinas; se destaca el ácido indol acético que es sintetizado a partir de triptófano que puede derivarse de las vías: indol-3-acetronilo, indole-3-acetamina, ácido indol-3-piruvico o de triptamina(48,50). Esta hormona es producida por algunos diazótrofos, por ejemplo: Stenotrophomona spp., Pseudomona spp(49), Azospirillum spp(51), Azotobacter spp. y Pseudomonas spp(26). Su principal efecto se relaciona con la modificación de la estructura, elongación e incremento de biomasa de raíz del forraje(37), lo que se favorece la absorción de nutrientes.
El estímulo hormonal que puede causar indirectamente la aplicación de diazótrofos en la planta puede favorecer su plasticidad fenotípica en ambientes sombreados(23), en condiciones de sequía(15) o suelos salinos(46). Fisiológicamente, la tolerancia a condiciones de estrés se relaciona con un incremento en la actividad de las enzimas superóxido-dismutasa y catalasa que eliminan el H de los radicales libres generados en condiciones estresantes(32). También, se reporta incremento en los contenidos de prolina, glutatión-reductasa(42), y en ACC-deaminasa(46).
Por otro lado, mayor disponibilidad de N en el suelo por efecto bacteriano, permite que la planta pueda aumentar la producción de clorofila al constituir parte de su estructura química, lo que conlleva a un incremento en la tasa fotosintética de la planta y en consecuencia en la producción de biomasa(32). Composicionalmente, puede favorecer el contenido de proteína cruda del forraje(1) y la producción de ácidos grasos insaturados(14).
Pese a los sinergismos mencionados, se reportan repuestas antagónicas con la inoculación de diazótrofos(2), por efecto de la inactivación de la nitrogenasa por la exposición a altas dosis de N. Aunque la falta de respuesta también puede deberse a una baja dosis de inoculante aplicado(23), que puede ser inhibido por control alelopático de la planta, lo que genera baja supervivencia, adaptación, y persistencia de los microrganismos inoculados. Incluso, la variabilidad entre el ecosistema puede limitar la respuesta de las bacterias debido a que el proceso de FBN se da solo en ambientes favorables que permiten la persistencia del grupo taxonómico de alfa-proteobacterias(9, 51).
Conclusiones
La FBN es la principal fuente de N en praderas perennes donde no se aplica N de origen sintético y en áreas de fuerte sequía, donde la planta logra mantener su crecimiento gracias a adaptaciones estructurales como la reducción del material aéreo para incrementar la longitud radicular. Se desconocen los mecanismos específicos de señalización que permiten la expresión de proteínas para la producción de hormonas y enzimas que hacen posibles dichas modificaciones y potencializan comunidades microbianas especializadas en la FBN para favorecer la supervivencia de la planta a condiciones extremas. Sin embargo, se ha identificado que las especies Brachiaria spp. y Pennisetum spp. tienen alto potencial en contribuir con el proceso de FBN, por la persistencia de alfa proteobacterias en la rizosfera y en el tejido de raíces, tallos y hojas.
Se destaca Azospirillum spp. y Azotobacter spp., pero de estos, Azospirillum brasilense tiene mayor potencial en fijar N por la capacidad de infectar tejido del forraje, lo que facilita eventualmente su supervivencia. Sin embargo, se desconoce si la colonización de este aislado junto con otros microrganismos endófitos resiste el sistema de defensa de la planta durante tiempos de exposición prolongados, y quizás esto, se relacione con la falta de respuesta productiva con la aplicación de algunos inoculantes. Es por esto que el desarrollo biotecnológico de estos productos apunta al estudio de microrganismos nativos, para evitar una respuesta alelopática negativa por la planta.
El incremento de materia seca con la aplicación de biofertilizantes es la principal respuesta observada según el análisis de revisión, este efecto puede permitir eventualmente intervalos entre pastoreos más cortos e intensificar las rotaciones en los sistemas ganaderos. También se ha observado que la aplicación de diazótrofos puede estimular la plasticidad fenotípica de la planta en condiciones de sombra, razón por la que el uso de biofertilizantes puede ser una opción rentable en sistemas silvopastoriles.
Aún existen desafíos como garantizar interacciones positivas entre los microorganismos aplicados y las cepas nativas; desarrollar biofertilizantes combinados con fertilizantes químicos y bioestimulantes; reducir los costos técnicos de aislamiento, masificación y obtención del producto final; formular productos por cultivo y de acuerdo con la etapa de crecimiento; utilizar métodos de seguimiento para la detección y cuantificación de poblaciones bacterianas persistentes que permitan ajustar la dosificación y frecuencia de uso de los biofertilizantes de acuerdo al manejo, cultivo, condiciones ambientales y tipo de suelo; y fomentar su aplicación en los sistemas de granja como servicio eco sistémico.
Agradecimientos y conflicto de interés
A Minciencias convocatoria 779 de 2017, regalías de la Gobernación del departamento de Boyacá – Colombia por el apoyo financiero y Universidad del Cauca proyecto ID. 5150 por el apoyo técnico, además a la Universidad Pedagógica y Tecnológica de Colombia UPTC y al Grupo de Investigación En Bioquímica y Nutrición Animal GIBNA.
Los autores no tienen conflicto de interés.
Literatura citada: