https://doi.org/10.22319/rmcp.v15i3.6479
Review
Winemaking by-products and grape polyphenols extracts as phytogenic feed additives in the pork production. Review
María Alejandra Ospina-Romero a
Humberto González-Ríos a*
Miguel Ángel Barrera-Silva b
Martin Valenzuela-Melendres a
Miguel Ángel Martínez-Téllez a
Araceli Pinelli-Saavedra a
a Centro de Investigación en Alimentación y Desarrollo, A.C. Carretera Gustavo Enrique Astiazarán Rosas, No. 46, Col. La Victoria, 83304, Hermosillo, Sonora, México.
b Universidad de Sonora. Departamento de Agricultura y Ganadería, Sonora, México.
*Corresponding author: hugory@ciad.mx
Abstract:
Winemaking by-products (WBP) such as grape pomace (GP), seed, and skin represent a rich source of phenolic compounds (PCs), fiber, fatty acids, and essential nutrients. Due to their profile and content of PCs, WBP can exert multiple bioactive properties on animal health, nutrition, and production. Specifically, beneficial effects have been observed in pigs. Therefore, these by-products and wine polyphenols extracts have been considered as valuable ingredients and as promising alternative to replace conventional resources of monogastric diets and thus minimize feeding cost. Nevertheless, these by-products are discarded and improperly disposed. Indeed, only 3% of the recovered by-products are used in animal nutrition without prior treatment. Emphasis has been placed on generating added value to obtain more significant economic and technological benefits and greater efficiency in animal production. This review discusses the most relevant and recent studies on the inclusion of WBP and their PCs during different stages of the pork production system (gestation-lactation, weaning, growth, and finishing) and their effects on the final quality of pork products. Additionally, strategies and treatments applied for the use of pomace in pig diets are described.
Keywords: Bioactivity, Phenolic compounds, Grape pomace, Monogastric, Phytochemicals.
Received: 03/06/2023
Accepted: 27/11/2023
Introduction
In view of the continuous intensification of pig production systems to meet world demand for animal products, the cultivation of grains (corn, soybeans, sorghum) has been frequently used as energy and protein sources for monogastric feed(1). In this context, it is estimated that 60 % of the biomass generated for feed production is used by the livestock sector and this item represents up to 70 % of the total costs of a swine farm(2). This dependence on agricultural crops has promoted food-feed competition between human consumption, the livestock sector and biodiesel production(1-3). Therefore, to face this problem, various economic, viable and promising alternatives have been sought, through which the use of conventional dietary ingredients can be minimized without affecting their nutritional contribution, and at the same time improve productivity, animal health and the final quality of animal foods(1,3). In this sense, an attractive option could be the inclusion of agro-industrial by-products and their extracts as phytogenic feed additive in monogastric nutrition(3–5). These wastes are generated in large amounts each year (1.3 billion tons) and represent a rich source of bioactive compounds (BCs). However, their use has been inefficient due to their low economic value that lacks alternative uses, and therefore most of these residues tend to be wasted and disposed of inadequately, generating a negative environmental impact(6). A significant amount of these by-products come from the winemaking process for which 75 % of the grapes harvested are used(7,8) while the remaining (25 %) represents the by-products (residues of skin, pulp, stems, and seeds) obtained during the pressing process (called pomace) and are discarded by the wine industry (13.1 million tons)(7–11). Inclusive, it is estimated that approximately 70 % of the phenolic content is retained in the pomace after processing. In this context, pomace and seeds are considered a rich source of PCs (tannins, anthocyanins, phenolic acids, and resveratrol); and possesses multiple bioactive properties that contribute to health and animal production(12,13). Although approximately 30 to 40 % of WBP have been valued worldwide (5.24 million tons) for use in the agricultural sector(13), it is estimated that only 3 % of the pomace produced is reused for animal feeds (0.39 million tons) without prior treatment(8,14). Therefore, in recent years, there has been an emphasis on generating added value to WBP to obtain more significant economic and technological benefits and greater efficiency in animal production. Likewise, the use of agro-industrial by-products could represent a non-conventional feeding alternative for monogastric animals and as a circular economy strategy that promotes the sustainability of pork production systems(15). Thereby, WBP represent an excellent opportunity to recover multiple phytochemical substances as PCs(8,14,16) which present great potential as phytogenic additives in pork production considering the importance of this young class of feed additives for pig farmers(5,17,18). Interest in these natural additives has increased in recent decades in response to the ban on the use of antibiotics and beta-adrenergic compounds(19).
Scientific reports show some beneficial effects in pigs on a diet including different dosages of grape by-products and an inclusion limit of 9 % has been reported(3,20). However, there is variability and inconsistency in the results obtained. The studies about WBP and grape polyphenols extracts in production cycle of swine are limited, and it is necessary to highlight what advances have been achieved in each stage of the pork production cycle, to determine future areas of opportunity in animal nutrition research, due to the importance of each of these phases for the success of the pork production. Therefore, the purpose of this review is to discuss the most relevant and recent studies on the inclusion of WBP and their PCs during different stages of the pork production system (gestation-lactation, weaning, growth, and finishing) and their effects on the final quality of pork products. Emphasis has been placed on the strategies and treatments applied to GP for inclusion in monogastric diet. Additionally, it is essential to evaluate the beneficial effects of including pomace phytochemicals or the raw matrix on productive efficiency.
Importance of winemaking by-products as alternative resources and feed additive in animal feeding
Grape is one of the most important fruit crops worldwide (7.3 million hectares), and about 36 % of total production (72.9 million tons) is concentrated in China, United States, and Italy(7,8,21). It is estimated that 52 % of this grape volume is pressed and destined for wine production (34.1 million tons)(22). During the final fermentation process, valuable by-products are obtained with multiple technological and health benefits, representing approximately 25 % of the total volume of grapes required by the wine industry(10). Vinification residues are classified into two categories: solids (stems, pomace, and seeds) and liquids; the former is generated during the harvesting and pressing of the grapes, whereas the latter is obtained during winemaking(23). The solid residues consist of 45 % pomace, 7.5 % stems, 6 % seeds, and other residues(24). The pomace represents the leading organic waste or by-product generated during the process of separating the solid waste (skins, seeds, and stems) from the liquid product or juice and constitutes 20 to 30 % of the processed grape(10). These residues are a rich source of various nutrients (protein, fatty acids, fiber) and PCs. However, these by-products are discarded, and their proper disposal represents one of the most critical challenges facing the wine industry.
Physicochemical composition of grape pomace
The composition of GP is determined by intrinsic factors such as grape maturity, variety, and sanitary conditions. Additionally, factors such as viticultural practices, edaphoclimatic conditions, harvest practices, and winemaking processes are determinants of composition. Fresh GP represents a rich source of dietary fiber (DF), that is composed of non-starch polysaccharides, tannins, and lignin(25,26). The fiber proportion in the pomace ranges from 43 to 7 5%. There are significant differences in soluble fractions, the insoluble fraction represents 61.3 to 73.5 %, while the soluble fraction compromises 10 and 3.7 % respectively. It is concentrated in the seeds (Table 1) and is higher in red varieties (51 %) than in white varieties (28 %)(13). Due to its high fiber content, pomace has been used as a feed additive in the diets of sows and the finishing stage; however, low inclusion rates have been reported in monogastrics (3 %), although other authors have obtained satisfactory results with rates of 9 %(3,20,27). Satisfactory results have been obtained regarding the production of volatile fatty acids (VFA) and beneficial bacteria(27). This food matrix also contains phenolic fractions that give it antioxidant properties and greater complexity(21). It is estimated that the content of PCs oscillates around 0.9 %, and the proportion of condensed tannins is 17.43 % and 7.29 % for red and white GP, respectively. However, the fiber and condensed tannin contents in pomaces could limit the bioavailability of dietary nutrients, causing a reduction in diet quality and therefore in the productive performance of animals(21,28,29). Bioavailability is determined by the inclusion of this by-product in the diet and the previous treatment(9). The moisture content of this by-product can range from 55 to 75 %, depending on processing conditions and is considered an indicator of the microbial and enzymatic degradation of fresh GP(30); this is one of the limitations of its application. Although low counts of aerobic mesophiles (3-6 Log CFU/g) and molds and yeasts (3 to 6 log CFU/g) have been reported, few studies have been conducted on the microbiological quality of this by-product(31,32). The protein content varies from 6 to 15 % of dry matter and is slightly higher in the skin than in the seeds(33). GP is rich in aspartic and glutamic acid, whereas it is deficient in sulfur amino acids and tryptophan. Additionally, it has been reported that unlike the seeds, the skin is a rich source of lysine and alanine. The lipid content of pomace derives from the seeds and ranges from 14 to 17 %. The lipid fraction is rich in unsaturated fatty acids and poor in saturated fatty acids. The fatty acids that predominate in grape seeds are linoleic (70 %), oleic (15 %), and palmitic (7 %). Likewise, it is necessary to highlight that its nutritional value is close to that of other foods such as corn and it has been reported that its crude protein content can be like that of conventional flours, which emphasizes its potential as a functional ingredient in animal feed(3).
Table 1: Chemical composition (g/100 g D.M) and phenolics compounds content of grape pomace, skin, and seeds (g /kg as D.M.)
Phenolic composition of grape pomace
WBP such as GP are considered a rich source of PCs, and these have been shown to possess multiple bioactive properties on animal health and production: antioxidant, antimicrobial, immunomodulator, anti-inflammatory(34). These phytochemicals are secondary metabolites of plants that act as a defense mechanism against pests, pathogens, herbivores, environmental factors, and stressful situations(35,36). Their chemical structure determines their biological properties and, therefore, their beneficial effects on animal health. A PC has one or two aromatic rings with one or more hydroxyl group substituents; this conformation determines its ability to capture free radicals(9). These compounds are classified into four categories based on their chemical structure and molecular weight: hydroxycinnamic acids (HA), stilbenes, lignans, and flavonoids(33). The latter is the broadest and most diverse group of polyphenols and has been studied most frequently in animal nutrition(37,38).
The main compounds found in GP are flavanols (catechin, epicatechin, and epigallocatechin), anthocyanins (cyanidin, pelargonidin, and delphinidin), and condensed tannins (Table 1). The former predominates in white grapes, whereas the latter is only present in red grapes(39). Grape skins and seeds are rich in epigallocatechin and gallocatechin, whereas anthocyanins and stilbenes such as resveratrol are found in the skin. Hydroxycinnamic acids predominate in the form of tartaric esters (caftaric, and courtaric) both in the skin and in the pulp of the grape, whereas the seeds are rich in gallic acid and protocatechuic acid. However, the phenolic composition varies depending on climatic conditions, growth, and fermentation time(9).
The beneficial effects attributed to these PCs include antimicrobial, antioxidant, anti-inflammatory, antimicrobial, immunomodulatory, cardioprotective, antidiabetic, anthelmintic, and intestinal microbiota modulation properties(36). The antioxidant activity exerted by PCs, mainly resveratrol (RES), in production animals has been emphasized. In this context, it has been suggested that the antioxidant capacity of RES is more significant than that provided by vitamins C and E; i.e., it is more effective in capturing and preventing free radicals(40,41). In turn, the PCs present in GP are readily accepted by consumers and producers, as pomace has been considered an alternative to traditional medicine for decades(41).
Potential use of winemaking by-products (grape pomace, seeds, skin) and its extracts as phytogenic feed additive in swine nutrition
In the last decade, agro-industrial by-products such as GP have received significant attention as alternative sources of unconventional animal feed(18). This by-product has a unique polyphenol profile(42). The content and diversity of PCs have stimulated in this food matrix as a supplement or additive to monogastric diets. Recently, these phytochemicals have been used as additives in feeds for pigs and poultry through various inclusion strategies (whole portions of the plant resource, by-products, extracts, isolated BCs, and complex blends of compounds). It is estimated that approximated 70 to 80 % of the industry that manufactures feed for monogastric uses phytochemicals as additives(36). The use of heterogeneous blends of these secondary metabolites with different plant origins (phytogenic) has been emphasized, whereas studies on the isolated forms of the compounds have been limited.
Phytogenic additives refer to compounds of plant origin that have been used in monogastric feed to improve animal productivity, health, and feed quality. These food additives have been classified into four categories according to their functionality in feed or/and animals: 1) Sensory (modifying the palatability and organoleptic properties of the feed); 2) Technological (acting as mycotoxin sequestrants and antioxidants); 3) Zootechnical (acting as immunomodulators, growth promoters of nonmicrobial origin, intestinal function modulators, digestive stimulants, or enhancers of quality in animal products, or productive and reproductive performance); and 4) Nutritional (acting as vitamins, minerals, or enzymes). Within these categories, they have been used as technological and zootechnical additives in pork production, particularly during the gestation-lactation and weaning stages, respectively(40,41) .
The dietary inclusion of phytochemicals and phytogenics in monogastric has shown that these compounds exert multiple beneficial effects on the animal due to their bioactive properties(43). The benefits including, improvements in palatability, productive performance (feed intake, average daily gain, and feed conversion), carcass quality, blood profile, animal welfare, health (immunity, antioxidant, antimicrobial), intestinal function (gastrointestinal morphology and nutrient digestibility) and as growth promoter(36,40). However, the variability among reports regarding the efficacy of some BCs subjected to specialized treatments and the partial understanding of their possible mechanisms of action have limited their use as feed additives. Therefore, future applications will be determined by the characteristics of the vegetal resource (primary and secondary compounds), complete knowledge of the mechanism of action, and safety of both the animals and the products generated(17).
Potential use of winemaking by-products in feeding pigs at different production stages
Impact on the function and reproductive organs of boars and sows
Reproduction represents a fundamental and integral component of sustainable swine production systems(42,44). It is also a complex process that greatly influences nutritional and feeding factors. The intensive inclusion of polyphenols in the diets of breeding animals has been evaluated(42). Typically, these compounds are obtained from alternative food sources such as agro-industrial by-products and natural feeding systems and have been used as additives to enrich animal diets(45). Previous research has evaluated the bioavailability of PCs in reproductive organs (ovary, uterus, placenta, fetus, and testis), the pituitary gland, and the hypothalamus and has shown that these PCs can cross diverse blood barriers and exert their physiological effects in the animal(46). However, the type of polyphenols, the selectivity of the tissues, and the physiological status of the animal determine the bioavailability of PCs in reproductive organs and, therefore, their effect on different reproductive events.
In this context, including these compounds in the diet can generate positive or negative changes in events such as fetal programming, pregnancy, gametogenesis, sexual behavior, reproductive function, and hormonal secretion in boars and sows. These effects are attributed to similarities in the chemical structure of PCs and estrogens; as a result, they can activate estrogen receptors (ERs) to exert a hormone-like effect that can modify the hormonal balance and reproductive events mentioned previously(42). Additionally, these compounds can control the expression of genes and the activity of sexual enzymes involved in regulating certain reproductive phases(47). Recently, the effects of PCs such as resveratrol and epigallocatechin have been evaluated in various assisted reproductive techniques (ARTs) in males and females (Figure 1). Thus far, promising results have been obtained during the thawing and cryopreservation of boar semen. It has been reported that the inclusion of epigallocatechin 3-gallato (EGCG) (25, and 50 µM) increases the fertilization rate of ovules, while the inclusion of resveratrol (0, 0.5, 1, and 2 mM/mL) and GP (2 and 4 %) maintains the integrity of the acrosome, sperm viability and reduces lipid peroxidation of fresh and cooled semen(47,48). These results have been attributed to the antioxidant activity of the PCs. In contrast, other studies conducted on in vitro fertilization have shown that compounds such as EGCG (0, 5, and 50 g/mL) can inhibit steroidogenesis and proliferation of granulosa cells in sows. Similarly, a decrease in progesterone synthesis and the percentage of fertilized oocytes was observed when a dose of 25 µg/mL of EGCG was included(49). The effect of these phytochemicals on the reproductive cycle of farm animals is still controversial and inconsistent(42).
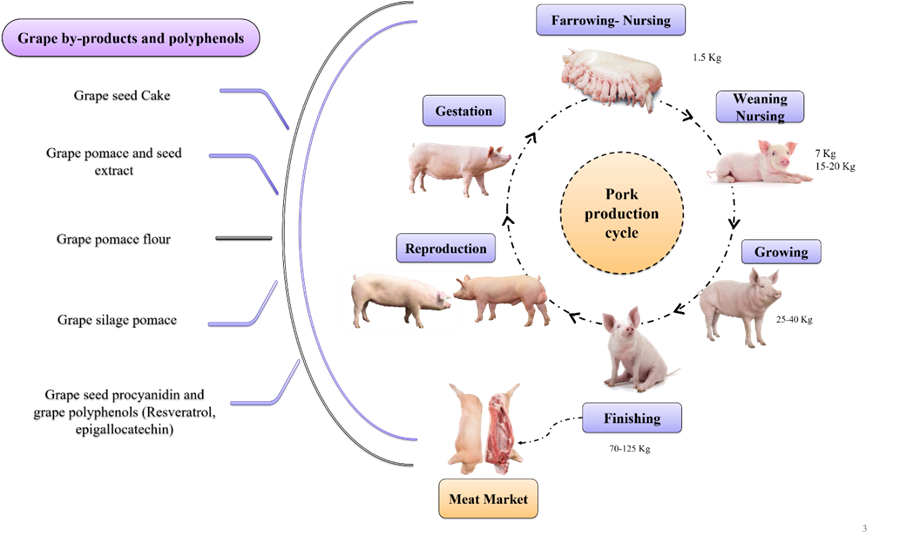
Figure 1: Grape by-products as phytogenic additives in pork production cycle
Gestation phase: Effects in pregnant sows, development of fetal piglets and farrowing
Oxidative stress represents one of the main critical points that swine production systems must face during the gestation (early, middle, and late) and lactation phases, in which systemic damage occurs in response to the high level of reactive oxygen species (ROS)(41,50). Gestation reflects a state of high oxidative stress in which various tissues and organs that actively metabolize oxygen (O2), such as the placenta, can present dynamic changes that modify the metabolic state of both the sow and the fetus. This state responds to an increase in O2 consumption and energy levels required to support the metabolic load involved in placental, embryonic development and fetal growth, which implies tissue mobilization. However, an excess of ROS impairs the physiological function of the placenta, triggering uterine restrictions that increase embryonic mortality rates and impair the development of the progeny(50).
Therefore, to mitigate the impact of oxidative stress, different antioxidant sources have been included in the diets of gestating sows. In this context, previous studies(50) have determined that the inclusion of 200 mg/kg and 300 mg/kg of grape seed polyphenols (GSP) in multiparous sows during late gestation (d 80) reduced stillbirth (0.63) and increased farrowing survival (89.32 %). These results reflected higher circulating levels of progesterone and estrogen in serum of pregnant sows (d 110), which are associated with pregnancy maintenance and maternal recognition(50). Likewise, this supplementation showed a significant improvement in antioxidant status of pregnant sows by increasing the enzymatic activity of superoxide dismutase (SOD) and glutathione peroxidase (GSHpx)(41,50). Similarly, supplementing of resveratrol (300 mg/kg) y catechins (200 and 300 mg/kg) from early (d 20) and mid gestation (d 40) until farrowing improved the antioxidant status (SOD, GSHpx, and catalase [CAT]) of sows and, their progeny during lactation phase(51,52). These results were attributed to the Keap 1-Nrf2 and Sirt 1 pathways regulating placental antioxidant genes(41). This same behavior has also been reported with the inclusion of other plant extracts, which reaffirms the importance of adequate maternal supplementation from any third of gestation to ensure adequate performance at parturition and during lactation(53).
Lactation phase: Lactating sows and suckling piglets
Like late gestation and parturition, lactation also leads to an overproduction of ROS, triggering lipid peroxidation. In this case, oxidative stress is attributed to the heightened metabolic energy demand required for mammary gland development and milk production. Moreover, the severity of oxidative damage affects the reproductive performance of the sow and hampers the adequate development of the offspring during the early stages of life. Therefore, maternal supplementation with phenolic compounds from gestation through lactation has been evaluated as a strategy to enhance the antioxidant status in lactating sows, which in turn may lead to improved development, pre-weaning performance, and overall health of their progeny (suckling piglets)(53).
In this context, it has been determined that the inclusion of GSP (200 and 300 mg/kg), resveratrol (200 and 300 mg/kg), and standardized blend of polyphenols-Proviox (catechin, procyanidins, and anthocyanins) increases the content of immunoglobulins (IgG and IgM) in the colostrum of supplemented sows(50), thereby improving the antioxidant status of suckling piglets by enhancing the enzymatic activity of CAT, SOD, and GPx(41,54,55). In addition, preweaning survival rate improved (96.9 %) with GSP supplementation (200 and 300 mg/kg) while the inclusion of resveratrol in the sows’ diets increased litter weight (57.26 kg vs 48.98 kg) as did the weight of the piglets at weaning (5.84 kg vs 5.24 kg), in comparison to unsupplemented females(41,50). Similarly, with the inclusion Proviox, a significant effect was observed in litter weight of sucking piglets and colostrum(54). Therefore, the nutritional strategies that have been evaluated at this stage are focused on improving the antioxidant status of pregnant sows and thus attenuating the impacts of oxidative stress, which, can be associated with an improvement in immune status and this information suggests that is possible to obtain higher number of piglets for the growing and finishing phases.
Weaning and postweaning phases
The weaning stage represents a change in piglet diet (from liquid to solid) and separation from the dam. Moreover, the establishment of social hierarchies, triggers a series of physiological responses that compromise immune function, intestinal metabolism, and antioxidant capacity, favoring the appearance of oxidative stress and an increase in the rates of morbidity and mortality(56). At the same time, feed intake decreases, causing growth retardation. The addition of WBP has been evaluated as a mitigating factor for this situation and as an alternative to replace the use of antibiotics as growth promoters in this phase(19). In this context, it has been demonstrated that the addition of PCs such as tannic acid (500 mg/kg and 1000 mg/kg) in the diets of weaned piglets (3-wk-old for 14 d) has beneficial effects on the transport of nutrients at the intestinal level along with the intestinal microbiota and morphology(19,57). This reflects the potential of GP as a source of various PCs. A previous study determined that the inclusion of 5 % dried GP improved the antioxidant status of the liver, kidney, and spleen in weaned piglets. Reduction in lipid peroxidation (MDA) and triglycerides was observed on d 36, whereas CAT, GSHpx, and SOD activities increased(58). Similar effects on enzyme activity and total antioxidant capacity (T-AOC) have been reported by other authors as presented in Table 2.
A recent study(59) reported that the inclusion of grape seed flour (8 %) in starter diets attenuates the oxidative stress induced by aflatoxin B1 (AFB1) and decreases inflammation markers (Table 2). In this context, a protective effect against aflatoxicosis has been shown, and the antioxidant status of birds improved with resveratrol supplementation (0.5 and 1 %)(58). Other studies have reported that agroindustrial by-products such as white GP (Malvasia) and red GP (Primitivo) have an excellent ability to adsorb ochratoxin (OTA), zearalenone (ZEN), fumonisin (FB1), and AFB1from aqueous solutions at pH 3 and 7(60). These results show that these compounds have considerable potential to sequester mycotoxins in weaned piglet diets. The study showed that the inclusion of white pomace (Malvasia 2.8%) in the diet of weaned pigs reduces gastrointestinal absorption of the mycotoxins ZEN and OTA by 67 and 69 %, respectively. Although the mechanism of action as a mycotoxin sequestrant has not yet been elucidated, it is suggested that the cellulose content in the pomace has a considerable potential to absorb AFB1 through electrostatic attractions; polyphenols may also could form complexes with mycotoxins(61).
Other biological activities, such as immunomodulatory, anti-inflammatory, and intestinal modulation have been evaluated more frequently in weaned piglets, given their immature digestive organs and immune systems. Evaluation has been emphasized in the first 3 wk after weaning because the immune system can only generate an effective response at 5 or 6 wk of age(62,63). In this context, it has been shown that supplementation with 50,100, and 150 mg/kg grape seed procyanidins(46); 1 % grape seed and grape marc extract (GSGME) with a phenol content of 8.5 % (piglets 7 kg BW for 4 wk)(49,64); and 5 % dried GP (20.41 mg/g dry matter, 5-wk-old for 28 d) decreases the incidence of diarrhea(58,62) increases the height of intestinal villus: depth ratio of crypts in the duodenum (VCR)(62) and jejunum(65); increases the proportion of Olsenella umbonata, Lactobacillus delbrueckii, and Selenomonas bovis in the cecum(65); decreases the populations of Streptococcus and Clostridium; and decreases VFA levels in the fecal microbiota of weaned piglets (5-wk-old for 25 d)(58) .
These results were attributed to the potential of PCs to improve antioxidant status, reduce intestinal permeability, increase surface area, improve nutrient absorption, and modulate intestinal populations by increasing butyrate-producing species that enhance colon health. These beneficial effects could be related to microbial metabolites such as 4-hydroxyphenylvaleric, 3-hydroibenzoic, caffeic, syringic, and protocatechuic acid(66). Similarly, other studies have verified that the PCs of WBP reduce the expression of proinflammatory genes in the intestine (cecum, ileum, and colon)(67). Supplementation with GSF (grape seed flour) in weaned piglets in an induced colitis model (iron dextran sulfate) attenuated the effect of this inducer, modulated the colonic microbiota, and reduced the impact of intestinal dysbiosis(25,27). Therefore, the PCs of the WBP represents a strategy to inhibit inflammation and modulate intestinal health during the postweaning stage. Among the productive parameters, a significant improvement of 4 to 7 % was observed in the weight gain:feed ratio (G:F)(67), and it has been suggested that this is due to an improvement in nutrient digestibility(25). However, other authors did not obtain a significant improvement in this indicator and stated that there was no consistent effect on intestinal morphology(68). Therefore, the improvement in G:F has been associated with reduced expression of proinflammatory genes and changes in microbial composition.
Growth and finishing stages
Previous studies have shown that grape seed pomace cakes (GSC 5% of the basal diet), as a rich source of proanthocyanidins, stilbenes, and flavanols (catechins, epicatechins, gallocatechins, epigallocatechins, and procyanidins), increased IgA levels (49.9 %) in animal plasma(41). Additionally, pomace decreased the inflammatory response in the liver and spleen, together with the production of hepatic cytokines (IFN-γ, IL-1, IL-8, and IL-6), and the gene expression and concentration of proinflammatory markers (IL-1β and, IFN-γ) in the spleen(63). Similarly, with the same dose, it was reported that cholesterol levels were reduced, which is attributed to the ability of resveratrol and epigallocatechin to bind to key regulators of liver lipid metabolism(69). Although malondialdehyde (MDA) levels decreased in the liver (13 %), the results were inconsistent in terms of enzyme gene expression; CAT, SOD, and GPx activities, T-AOC; which indicates that the antioxidant status of the pigs did not improve during supplementation and can be attributed to the low CF content in the matrix. In contrast, other authors(63,68) reported a significant increase in the expression of enzymatic genes (CAT, GPx); a similar trend in the activity of CAT, SOD, GPx, and T-AOC (49 %); and decreased MDA levels in the spleen (20 %). Similarly, with the addition of 9 % GP silage during the growth stage, the decomposition of H2O2 increased(20).
It has been indicated that the inclusion of 1 % GSGME with a polyphenol content of 8.5 % and the addition of 3 % fermented GP is effective in improving the productive performance (body weight, average daily gain [ADG], intake feed, and feed conversion) in growing pigs(70). However, it has also been shown that the inclusion of GSC (5 %) in finisher pigs does not influence the productive performance(64). Similar results were obtained by other authors in all finishing stage(62). Although GP has been reported to improve sensory abilities and metabolism in pigs(58), few specific studies for the finishing stage (> 70 kg) have evaluated any by-product of vinification, unlike the first two production phases(71). In general, the investigations integrate the initiation phase with growth(27). Even the results of GP supplementation on productive performance (ADG, feed conversion, and feed intake) are inconsistent(17) and it has been suggested that this nutritional matrix may suppress growth performance. The concentration and profile of PCs will determine this impact together with the binding to digestive enzymes and proteins(72).
A study on pigs reported that the inclusion of 300 and 600 mg/kg of resveratrol in finishing diets for 49 d induced a transition in the type of muscle fibers and a change in energy metabolism(73) that favored the expression of oxidative fibers (IIA) over glycolytic fibers (IIB). These results are consistent with those reported by other authors, who observed a lower proportion of IIB fibers when animals were supplemented with 400 ppm and 600 ppm of resveratrol for 42 d, which could indicate a growth-promoting effect(74). However, the authors did not observe a significant effect on production efficiency. Likewise, it has been shown that with the inclusion of dry pomace (5 %) in finisher pigs, the profile of polyphenols in plasma (273 nm and 279 nm) is modified and these changes could be reflected in the muscle(75).
Effects of winemaking by-products and wine polyphenols extracts in pork meat quality, nutritional value and meat products
Grape pomace supplementation of animal feed or its direct inclusion in meat or meat products exerts an antioxidant and antimicrobial effect that determines the final quality of the product. Likewise, it has been reported that the oxidative stability of meat products is determined by the composition of unsaturated fatty acids(76,77). Although a higher PUFA content in meat has been associated with greater susceptibility to lipid oxidation, it has also been shown that the inclusion of sources rich in PCs gives it greater stability, which is attributed to an increase in antioxidant enzyme activity and a decrease in MDA and TBARS in the muscle of monogastric(1,3). The antioxidant effects of pomace by-products such as grape seeds and skins, which have been seen in animal production, have been associated with end products of the degradation of low molecular weight PCs, which can be transferred to tissues and responsible compounds such as epicatechin, among others, have been detected(51,76).
Therefore, it has received significant attention as a natural preservative during meat storage to extend shelf life. Indeed, it has been shown that oral supplementation of flavonoids (quercetin, epicatechin, and catechin) in the diet increases the in vivo concentrations of vitamin E(78), which could explain the antioxidant effect of PCs on animal tissues. Additionally, the storage and deposition dynamics of vitamin E differ in monogastric animals. In pigs, the accumulation of this vitamin in response to supplementation with grape pomace is significant in fat, liver, and muscle, whereas, in birds there is more deposition in the liver and less deposition in the fat(79). In addition, it has been reported that the inclusion of 30 g of fermented pomace/kg in the diets of pigs improved the percentage of marbling (3.5 %), increased the a* (25 %) and b* (45 %) values of the meat, and reduced MDA levels by 53 %(27). Similarly, pomace modifies the polyunsaturated fatty acids (PUFA) content and the SFA/PUFA ratio, which has been attributed to the moderating effect of grape pomace on vitamin E in the intestine(78). Likewise, it has also been shown that the supply of 5 % grape pomace from the feed of finisher pigs increases the levels of omega 3 fatty acids (1.27 times), alpha-linoleic (1.35 times) and eicosapentaenoic acid (1.30 times), while it tends to decrease the n-6:n-3 ratio with respect to the control(75).
Furthermore, it has been shown that the b* value is mainly affected by the type of myoglobin in the muscle(27). However, the supplementation of 3 %, 5 %, 6 %, and 10 % grape pomace with a mixture of oils from the finishing stage did not affect the production of thiobarbituric acid reactive substances (TBARS) in the pork loin samples(79) but increased the a* value together with the color saturation index, which suggests a possible antioxidant effect.
For texture of the meat, satisfactory results have been obtained in shear force, which have been associated with an increase in pH values in response to supplementation with grape pomace extracts. In addition, it has been proposed that grape proanthocyanin may decrease collagen deposition by suppressing the expression of protein promoters involved in collagen synthesis (TGF-β)(80).
On the other hand, in the maturation of meat products, it has been determined that for every 100 g of raw meat, a powdered grape pomace (1L) mixture should be used at 4 °C for 72 h. Marinating the pork tenderloin with this mixture, the crude protein, crude fat, ash, pH, a* value, and b* value in the meat decreased, and its shear force and moisture content increased. During meat storage, the concentration of TBARS, volatile basic nitrogen, and the total microbial count decreased(80,81). These results could be attributed to the organic acids and PCs present in the WBP, which could inhibit the growth of microorganisms by reducing the activity of some enzymes and inhibiting the production of basic substances(82). Therefore, marinating with grape pomace powder improves some meat quality characteristics and stabilizes the pork loin during storage. Similar results were observed when adding two different grape pomace extracts to pork burgers(83), resulting in an inhibition in lipid oxidation, greater color stability, and improved acceptability with the HLIP extraction method (Table 3).
Table 3: Addition of winemaking by-products on pork meat and pork products
* Supplementation from animal feed
Technological considerations for the inclusion of winemaking by-products in monogastric diets
WBP such as grape pomace, have been subjected to various pre-treatments (endogen enzymes, cellulolytic enzymes, polyethylene glycol, and fermentation) to increase the bioavailability of their BCs especially of PCs and improve its nutritional value to favor their inclusion in animal feed. In this context, it has been demonstrated that the inclusion of enzyme complexes (tannases) and polyethylene glycol in monogastric diets can partially inactivate condensed grape pomace tannins, minimize the content of antinutritional factors and increase antioxidant and antimicrobial activity(28,84). Among these treatments, fermentation processes with yeasts, bacteria, and fungi have been emphasized(27). In this context, a systematic biotechnological approach was developed, through which the metabolic capacities of several species of fungi (Aspergillus, Rhizopus, and Trichoderma ssp.) were evaluated about the bioconversion of grape pomace and lees to feed for animals(85). In this study, the authors obtained an improvement in the protein content (5 to 26 %) and digestibility of the feed (25 to 50 %).
Similarly, the supplementation of 3 % fermented grape pomace with Saccharomyces boulardii in pigs during their growth-finishing stage increased the apparent digestibility of dry matter (79 %) and nitrogen (82.5 %) after 11 wk(27). These results suggest that the PCs of this matrix can stimulate intestinal fermentation and influence the production of specific microbial metabolites, which explains the improvement in growth. This strategy makes it feasible to provide rich protein sources and an optimal nutritional status to the animals(66). However, it has also been reported that with the inclusion of 20 % fermented grape pomace in finisher pigs, the digestibility of dry matter, crude protein, energy, and essential and nonessential amino acids was reduced with respect to the control(25). These results were attributed to the formation of complexes between some PCs and the food protein, which limited the action of digestive enzymes; thus, it is necessary to consider the inclusion levels for each production stage.
Other technological processes such as the elaboration of flours, powders, and cakes from WBP (seeds, pomace, skins) are indispensable in the formulation of diets for monogastric animals. Therefore, the optimum conditions of temperature, time and particle size have been evaluated and standardized to obtain better results in animal feed with respect to the raw by-product. GP has been subjected to a drying process with hot air flow at 65 °C for 4 d, 60 °C for 20 h(16,86), 50 °C for 12 h(87) and 60 °C until reaching a constant weight. Also, after grinding, particle sizes from 1 mm to 6 mm have been defined for poultry and swine respectively(67,70,86). Likewise, it is also important to define and specify the temperature/time conditions for the GP intended for pig feeding and to standardize the particle sizes depending on the production stage, which has made it possible to obtain better results in animal feeding concerning the crude by-products(8,20,70).
Conclusions and implications
A large number of completed studies have evaluated the antioxidant, anti-inflammatory, antimicrobial, and immunomodulatory properties of wine industry by-products, emphasizing weaned piglets and early growth stages. However, the effect on muscle fibers with the raw matrix has not been determined. In the final stage of production, it is still necessary to continue evaluating different presentations of this matrix and new inclusion levels and consider mortality rates as another indirect measure of productive performance. The inclusion of grape pomace in extract or flour form and its BCs in the pig diets at each productive phase is considered a low-cost antioxidant and antimicrobial source that exerts multiple benefits on the productive performance of pigs as well as the meat quality. Therefore, it represents a promising alternative for the animal nutrition sector that could minimize the use of synthetic antioxidant compounds, growth promoters such as antibiotics, and beta-adrenergic agents in the pork production system.
Conflict of interest
The authors declare that there are no conflicts of interest.
Literature cited:
Table 2: Effects of phenolic compounds from winemaking by-products in pork production systems
a) Type of additive: Z= zootechnical; T= technological; S= Sensory; PP= productive performance; DR= reproductive performance; T-AOC (Total antioxidant capacity).
b) Results: MDA (malondialdehyde); AOS (Antioxidant status); NR= not reported, MIF= modulation of intestinal function; ADC= apparent digestibility coefficient.