https://doi.org/10.22319/rmcp.v15i3.6479
Revisión bibliográfica
Subproductos de la vinificación y extractos de polifenoles de la uva como aditivos fitogénicos para raciones en la producción porcina. Revisión
María Alejandra Ospina-Romero a
Humberto González-Ríos a*
Miguel Ángel Barrera-Silva b
Martin Valenzuela-Melendres a
Miguel Ángel Martínez-Téllez a
Araceli Pinelli-Saavedra a
a Centro de Investigación en Alimentación y Desarrollo, A.C. Carretera Gustavo Enrique Astiazarán Rosas, No. 46, Col. La Victoria, 83304, Hermosillo, Sonora, México.
b Universidad de Sonora. Departamento de Agricultura y Ganadería, Sonora, México.
*Autor de correspondencia: hugory@ciad.mx
Resumen:
Los subproductos de vinificación, como el orujo, la semilla y la piel de la uva, representan una fuente rica en compuestos fenólicos (CF), fibra, ácidos grasos y nutrientes esenciales, la cual ha recibido gran interés en los sistemas de producción porcina. Con respecto a su perfil y contenido de CF, en estudios previos se ha demostrado que estos subproductos pueden ejercer múltiples propiedades bioactivas sobre la salud, nutrición y producción animal, incluso se han observado efectos beneficiosos en el ganado porcino. Asimismo, estos subproductos y los extractos de polifenoles del vino se han considerado ingredientes valiosos, y una alternativa prometedora para sustituir los recursos convencionales de las dietas de monogástricos y así reducir el costo de alimentación. Sin embargo, estos subproductos se desechan y eliminan de forma inadecuada. De hecho, sólo el 3 % de los subproductos recuperados se utilizan en nutrición animal sin tratamiento previo. Por ende, se ha hecho énfasis en generar valor agregado a los subproductos de la vinificación para obtener beneficios económicos y tecnológicos más significativos y una mayor eficiencia en la producción animal. Esta revisión discute los estudios más relevantes y recientes sobre la inclusión de subproductos de vinificación y sus CF durante cada etapa del sistema de producción porcina (gestación-lactancia, destete, crecimiento y finalización) y sus efectos sobre la calidad final de la carne y sus derivados. Además, se describen estrategias y tratamientos aplicados para el uso del orujo en dietas porcinas.
Palabras clave: Bioactividad, Compuestos fenólicos, Orujo de uva, Monogástricos Fitoquímicos.
Recibido: 03/06/2023
Aceptado: 27/11/2023
Introducción
En vista de la continua intensificación de los sistemas de producción porcina para satisfacer la demanda mundial de productos de origen animal, el cultivo de granos (maíz, soya, sorgo) ha sido utilizado con frecuencia como fuentes de energía y proteínas para la alimentación de monogástricos(1). En este contexto, se estima que el 60 % de la biomasa generada para la producción de dietas es utilizada por el sector ganadero y este rubro representa hasta el 70 % de los costos totales de una explotación porcina(2). Esta dependencia de los cultivos agrícolas ha promovido la competencia alimento-pienso entre el consumo humano, el sector ganadero y la producción de biodiesel(1-3). Por lo tanto, para enfrentar esta problemática, se han buscado diversas alternativas económicas, viables y prometedoras, a través de las cuales se puede minimizar el uso de ingredientes dietéticos convencionales sin afectar su contribución nutricional, y al mismo tiempo mejorar la productividad, la salud animal y la calidad final de los alimentos de origen animal(1,3). En este sentido, una opción atractiva podría ser la inclusión de subproductos agroindustriales y sus extractos como aditivo fitogénico para piensos en la nutrición de monogástricos(3-5). Estos residuos se generan en grandes cantidades cada año (1,300 millones de toneladas) y representan una rica fuente de compuestos bioactivos (CB). No obstante, su uso ha sido ineficiente debido a su bajo valor económico que carece de usos alternativos, por lo que la mayoría de estos residuos tienden a ser desechados y eliminados de manera inadecuada, generando un impacto ambiental negativo(6). Una cantidad importante de estos subproductos proviene del proceso de vinificación para el que se utiliza el 75 % de las uvas cosechadas(7,8), mientras que el resto (25 %) representa los subproductos (residuos de piel, pulpa, raspones y semillas) obtenidos durante el proceso de prensado (denominados orujo) y son desechados por la industria vitivinícola (13.1 millones de toneladas)(7-11). Inclusive, se estima que aproximadamente el 70 % del contenido fenólico se retiene en el orujo después del procesamiento. En este contexto, el orujo y las semillas se consideran una rica fuente de CF (taninos, antocianinas, ácidos fenólicos y resveratrol); y posee múltiples propiedades bioactivas que contribuyen a la salud y a la producción animal(12,13). A pesar de que aproximadamente entre el 30 % y el 40 % de los SPV han sido evaluados en todo el mundo (5.24 millones de toneladas) para su uso en el sector agrícola(13), se estima que solo el 3 % del orujo producido se reutiliza para piensos de animales (0.39 millones de toneladas) sin tratamiento previo(8,14). Por ello, en los últimos años, se ha hecho énfasis en generar valor agregado a los SPV para obtener beneficios económicos y tecnológicos más significativos y una mayor eficiencia en la producción animal. Asimismo, el uso de subproductos agroindustriales podría representar una alternativa de alimentación no convencional para animales monogástricos y una estrategia de economía circular que promueva la sostenibilidad de los sistemas de producción porcina(15). De esta manera, los SPV representan una excelente oportunidad para recuperar múltiples sustancias fitoquímicas como CF(8,14,16), los cuales presentan un gran potencial como aditivos fitogénicos en la producción porcina considerando la importancia de esta clase de aditivos para piensos para los productores porcinos(5,17,18). El interés por estos aditivos naturales ha aumentado en las últimas décadas en respuesta a la prohibición del uso de antibióticos y compuestos beta-adrenérgicos(19).
Reportes científicos muestran algunos efectos beneficiosos en cerdos con una dieta que incluye diferentes dosis de subproductos de la uva y se ha descrito un límite de inclusión del 9 %(3,20). No obstante, existe variabilidad e inconsistencia en los resultados obtenidos. Los estudios sobre SPV y extractos de polifenoles de la uva en el ciclo de producción de cerdos son limitados, y es necesario destacar qué avances se han logrado en cada etapa del ciclo productivo porcino, para determinar futuras áreas de oportunidad en la investigación en nutrición animal, debido a la importancia de cada una de estas fases para el éxito de la producción porcina. Por lo tanto, el propósito de esta revisión es discutir los estudios más relevantes y recientes sobre la inclusión de SPV y sus CF durante diferentes etapas del sistema de producción porcina (gestación-lactancia, destete, crecimiento y finalización) y sus efectos en la calidad final de los productos porcinos. Se ha hecho énfasis en las estrategias y tratamientos aplicados al OU para su inclusión en la dieta de monogástricos. Además, es fundamental evaluar los efectos beneficiosos de la inclusión de fitoquímicos de orujo o de la matriz cruda sobre la eficiencia productiva.
Importancia de los subproductos de la vinificación como recursos alternativos y aditivos para raciones en la alimentación animal
La uva es uno de los cultivos frutales más importantes a nivel mundial (7.3 millones de hectáreas), y alrededor del 36 % de la producción total (72.9 millones de toneladas) se concentra en China, Estados Unidos e Italia(7,8,21). Se estima que el 52 % de este volumen de uva se prensa y se destina a la producción de vino (34.1 millones de toneladas)(22). Durante el proceso de fermentación final, se obtienen valiosos subproductos con múltiples beneficios tecnológicos y de salud, los cuales representan aproximadamente el 25 % del volumen total de uvas requerido por la industria vitivinícola(10). Los residuos de la vinificación se clasifican en dos categorías: sólidos (raspones, orujo y semillas) y líquidos; el primero se genera durante la cosecha y el prensado de la uva, mientras que el segundo se obtiene durante la vinificación(23). Los residuos sólidos consisten en un 45 % de orujo, un 7.5 % de raspones, un 6 % de semillas y otros residuos(24). El orujo representa el principal residuo orgánico o subproducto generado durante el proceso de separación de los residuos sólidos (pieles, semillas y raspones) del producto líquido o jugo y constituye entre el 20 y el 30 % de la uva transformada(10). Estos residuos son una rica fuente de diversos nutrientes (proteínas, ácidos grasos, fibra) y CF. Sin embargo, estos subproductos se desechan y su adecuada eliminación representa uno de los desafíos más críticos que enfrenta la industria vitivinícola.
Composición fisicoquímica del orujo de uva
La composición del OU está determinada por factores intrínsecos como la madurez de la uva, la variedad y las condiciones sanitarias. Además, factores como las prácticas vitivinícolas, las condiciones edafoclimáticas, las prácticas de cosecha y los procesos de vinificación son determinantes de la composición. El OU fresco representa una rica fuente de fibra dietética (FD), que se compone de polisacáridos almidonados, taninos y lignina(25,26). La proporción de fibra en el orujo oscila entre el 43 y el 7.5 %. Existen diferencias significativas en las fracciones solubles; la fracción insoluble representa entre el 61.3 y el 73.5 %, mientras que la fracción soluble comprende el 10 y 3.7 % respectivamente. Se concentra en las semillas (Cuadro 1) y es mayor en las variedades rojas (51 %) que en las variedades blancas (28 %)(13). Debido a su alto contenido en fibra, el orujo se ha utilizado como aditivo para piensos en las dietas de las cerdas y en la etapa de finalización; no obstante, se han reportado bajas tasas de inclusión en monogástricos (3 %), aunque otros autores han obtenido resultados satisfactorios con tasas de 9 %(3,20,27).
Se han obtenido resultados satisfactorios en cuanto a la producción de ácidos grasos volátiles (AGV) y bacterias beneficiosas(27). Por lo tanto, se han reportado bajas tasas de inclusión en monogástricos (3 %), aunque otros autores han obtenido resultados satisfactorios con tasas de 9 % (3,20,27). Esta matriz alimentaria también contiene fracciones fenólicas que le confieren propiedades antioxidantes y mayor complejidad(21). Se estima que el contenido de CF oscila alrededor del 0.9 %, y la proporción de taninos condensados es de 17.43 % y 7.29 % para los OU rojos y blancos, respectivamente. Sin embargo, el contenido de fibra y taninos condensados en los orujos podría limitar la biodisponibilidad de los nutrientes dietéticos, provocando una reducción en la calidad de la dieta y, por lo tanto, en el desempeño productivo de los animales(21,28,29). La biodisponibilidad está determinada por la inclusión de este subproducto en la dieta y por el tratamiento previo(9). El contenido de humedad de este subproducto puede oscilar entre el 55 y el 75 %, dependiendo de las condiciones de procesamiento y se considera un indicador de la degradación microbiana y enzimática del OU fresco(30), siendo ésta una de las limitaciones de su aplicación. A pesar de que se han descrito recuentos bajos de mesófilos aeróbicos (de 3 a 6 Log UFC/g) y de mohos y levaduras (de 3 a 6 log UFC/g), se han realizado pocos estudios sobre la calidad microbiológica de este subproducto(31,32). El contenido de proteína varía del 6 al 15 % de la materia seca y es ligeramente superior en la piel que en las semillas(33). El OU es rico en ácido aspártico y glutámico, pero es deficiente en aminoácidos azufrados y triptófano. Además, se ha reportado que, a diferencia de las semillas, la piel es una rica fuente de lisina y alanina. El contenido lipídico del orujo procede de las semillas y oscila entre el 14 y el 17 %. La fracción lipídica es rica en ácidos grasos insaturados y pobre en ácidos grasos saturados. Los ácidos grasos que predominan en las semillas de uva son el linoleico (70 %), el oleico (15 %) y el palmítico (7 %). Asimismo, es necesario destacar que su valor nutricional es cercano al de otros alimentos como el maíz y se ha reportado que su contenido de proteína cruda puede ser similar al de las harinas convencionales, lo que enfatiza su potencial como ingrediente funcional en la alimentación animal(3).
Cuadro 1: Composición química (g/100 g MS) y contenido de compuestos fenólicos del orujo, piel y semillas de uva (g/kg como MS)
Composición fenólica del orujo de uva
Los SPV como el OU se consideran una fuente rica en CF, y se ha demostrado que poseen múltiples propiedades bioactivas sobre la salud y la producción animal: antioxidantes, antimicrobianas, inmunomoduladoras, antiinflamatorias(34). Estos fitoquímicos son metabolitos secundarios de las plantas que actúan como mecanismo de defensa contra plagas, patógenos, herbívoros, factores ambientales y situaciones estresantes(35,36). Su estructura química determina sus propiedades biológicas y, por tanto, sus efectos beneficiosos para la salud animal. Un CF tiene uno o dos anillos aromáticos con uno o más sustituyentes del grupo hidroxilo; esta conformación determina su capacidad para capturar radicales libres(9). Estos compuestos se clasifican en cuatro categorías según su estructura química y peso molecular: ácidos hidroxicinámicos (AH), estilbenos, lignanos y flavonoides(33). Este último es el grupo más amplio y diverso de polifenoles y ha sido estudiado con mayor frecuencia en nutrición animal(37,38).
Los principales compuestos que se encuentran en el OU son los flavanoles (catequina, epicatequina y epigalocatequina), las antocianinas (cianidina, pelargonidina y delfinidina) y los taninos condensados (Cuadro 1). Los primeros predominan en las uvas blancas, mientras que los segundos solo están presentes en las uvas rojas(39). La piel y las semillas de la uva son ricas en epigalocatequina y galocatequina, mientras que las antocianinas y los estilbenos como el resveratrol se encuentran en la piel. Los ácidos hidroxicinámicos predominan en forma de ésteres tartáricos (caftáricos y coutáricos) tanto en la piel como en la pulpa de la uva, mientras que las semillas son ricas en ácido gálico y ácido protocatecuico. Sin embargo, la composición fenólica varía dependiendo de las condiciones climáticas, el crecimiento y el tiempo de fermentación(9).
Los efectos benéficos atribuidos a estos CF incluyen propiedades antimicrobianas, antioxidantes, antiinflamatorias, antimicrobianas, inmunomoduladoras, cardioprotectoras, antidiabéticas, antihelmínticas y de modulación de la microbiota intestinal(36). Se ha enfatizado la actividad antioxidante ejercida por los CF, principalmente el resveratrol (RES), en animales de producción. En este contexto, se ha sugerido que la capacidad antioxidante del RES es más significativa que la proporcionada por las vitaminas C y E; es decir, es más eficaz en la captura y prevención de radicales libres(40,41). A su vez, los CF presentes en el OU son fácilmente aceptados por los consumidores y productores, ya que el orujo ha sido considerado una alternativa a la medicina tradicional durante décadas(41).
Uso potencial de los subproductos de la vinificación (orujo, semillas, piel de uva) y sus extractos como aditivo fitogénico para dietas en la nutrición porcina
En la última década, los subproductos agroindustriales como el OU han recibido una atención significativa como fuentes alternativas de alimento no convencional para animales(18). Este subproducto tiene un perfil polifenólico único(42). El contenido y diversidad de CF han fomentado esta matriz alimentaria como complemento o aditivo a dietas de monogástricos. Recientemente, estos fitoquímicos se han utilizado como aditivos en piensos para cerdos y aves de corral a través de diversas estrategias de inclusión (porciones enteras del recurso vegetal, subproductos, extractos, CB aislados y mezclas complejas de compuestos). Se estima que aproximadamente entre el 70 y el 80 % de la industria que elabora dietas para monogástricos utiliza fitoquímicos como aditivos(36). Se ha enfatizado el uso de mezclas heterogéneas de estos metabolitos secundarios con diferentes orígenes vegetales (fitogénicos), mientras que los estudios sobre las formas aisladas de los compuestos han sido limitados.
Los aditivos fitogénicos se refieren a compuestos de origen vegetal que se han utilizado en la alimentación de monogástricos para mejorar la productividad, la salud y la calidad de los piensos para animales. Estos aditivos alimentarios se han clasificado en cuatro categorías según su funcionalidad en piensos y animales: 1) Sensoriales (modificando la palatabilidad y las propiedades organolépticas del pienso); 2) Tecnológicos (actuando como secuestrantes de micotoxinas y antioxidantes); 3) Zootécnicos (actuando como inmunomoduladores, promotores del crecimiento de origen no microbiano, moduladores de la función intestinal, estimulantes digestivos o potenciadores de la calidad en productos animales, o del rendimiento productivo y reproductivo); y 4) Nutricionales (actuando como vitaminas, minerales o enzimas). Dentro de estas categorías, han sido utilizados como aditivos tecnológicos y zootécnicos en la producción porcina, particularmente durante las etapas de gestación-lactancia y destete, respectivamente(40,41).
La inclusión dietética de fitoquímicos y fitogénicos en monogástricos ha demostrado que estos compuestos ejercen múltiples efectos beneficiosos sobre el animal debido a sus propiedades bioactivas(43). Los beneficios incluyen mejoras en la palatabilidad, el rendimiento productivo (ingesta de alimento, ganancia media diaria y conversión alimenticia), la calidad de la canal, el perfil sanguíneo, el bienestar animal, la salud (inmunidad, antioxidante, antimicrobiano), la función intestinal (morfología gastrointestinal y digestibilidad de nutrientes) y como promotor del crecimiento(36,40). No obstante, la variabilidad entre los reportes con respecto a la eficacia de algunos CB sometidos a tratamientos especializados y la comprensión parcial de sus posibles mecanismos de acción han limitado su uso como aditivos para piensos. Por lo tanto, las aplicaciones futuras estarán determinadas por las características del recurso vegetal (compuestos primarios y secundarios), el conocimiento completo del mecanismo de acción y la seguridad tanto de los animales como de los productos generados(17).
Uso potencial de los subproductos de la vinificación en la alimentación de los cerdos en diferentes etapas de producción
Impacto en la función y los órganos reproductores de verracos y cerdas
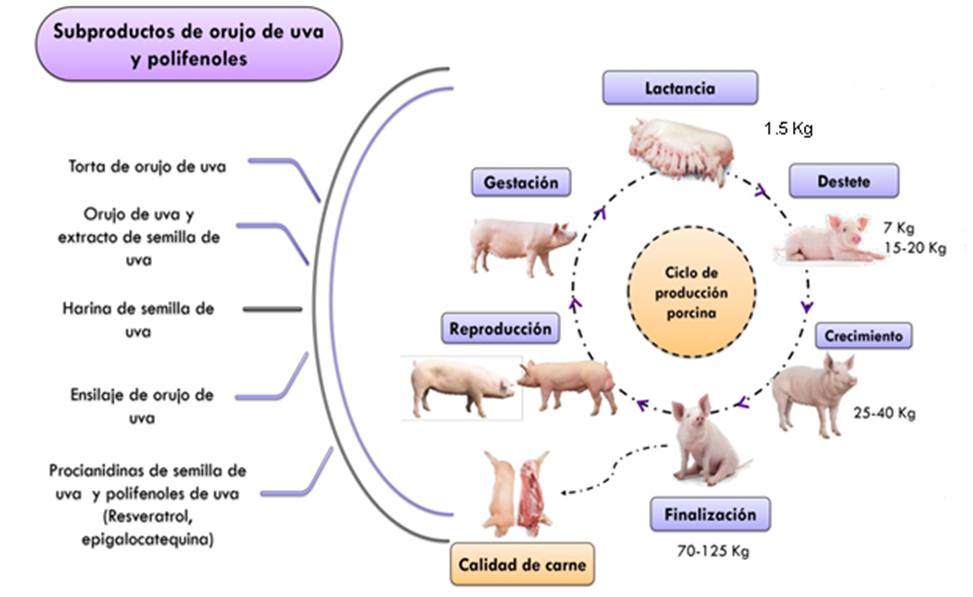
La reproducción representa un componente fundamental e integral de los sistemas sostenibles de producción porcina(42,44). También es un proceso complejo que influye en gran medida en los factores nutricionales y de alimentación. Se ha evaluado la inclusión intensiva de polifenoles en las dietas de animales reproductores(42). Típicamente, estos compuestos se obtienen de fuentes alternativas de alimentos, como subproductos agroindustriales y sistemas de alimentación naturales, y se han utilizado como aditivos para enriquecer las dietas de los animales(45). Investigaciones previas han evaluado la biodisponibilidad de los CF en los órganos reproductivos (ovario, útero, placenta, feto y testículos), la glándula pituitaria y el hipotálamo, y han demostrado que estos CF pueden atravesar diversas barreras sanguíneas y ejercer sus efectos fisiológicos en el animal(46). Sin embargo, el tipo de polifenoles, la selectividad de los tejidos y el estado fisiológico del animal determinan la biodisponibilidad de los CF en los órganos reproductivos y, por lo tanto, su efecto sobre diferentes eventos reproductivos.
En este contexto, la inclusión de estos compuestos en la dieta puede generar cambios positivos o negativos en eventos como la programación fetal, la gestación, la gametogénesis, el comportamiento sexual, la función reproductiva y la secreción hormonal en verracos y cerdas. Estos efectos se atribuyen a las similitudes en la estructura química de los CF y los estrógenos, por lo que pueden activar los receptores de estrógenos (RE) para ejercer un efecto similar al de las hormonas que puede modificar el equilibrio hormonal y los eventos reproductivos mencionados(42). Además, estos compuestos pueden controlar la expresión de genes y la actividad de las enzimas sexuales involucradas en la regulación de ciertas fases reproductivas(47). Recientemente, se han evaluado los efectos de CF como el resveratrol y la epigalocatequina en diversas técnicas de reproducción asistida (TRA) en machos y hembras (Figura 1). Hasta ahora, se han obtenido resultados prometedores durante la descongelación y criopreservación del semen de verraco. Se ha reportado que la inclusión de epigalocatequina 3-galato (EGCG) (25 y 50 μM) aumenta la tasa de fertilización de los óvulos, mientras que la inclusión de resveratrol (0, 0.5, 1 y 2 mM/mL) y OU (2 y 4 %) mantiene la integridad del acrosoma, la viabilidad de los espermatozoides y reduce la peroxidación lipídica del semen fresco y refrigerado(47,48). Estos resultados se han atribuido a la actividad antioxidante de los CF. Por el contrario, otros estudios realizados sobre la fecundación in vitro han demostrado que compuestos como la EGCG (0, 5 y 50 g/ml) pueden inhibir la esteroidogénesis y la proliferación de las células de la granulosa en las cerdas. Del mismo modo, se observó una disminución en la síntesis de progesterona y en el porcentaje de ovocitos fecundados cuando se incluyó una dosis de 25 μg/ml de EGCG(49). El efecto de estos fitoquímicos en el ciclo reproductivo de los animales de granja sigue siendo controvertido e inconsistente(42).
Figura 1: Subproductos de la uva como aditivos fitogénicos en el ciclo de producción porcina
Fase de gestación: efectos en cerdas gestantes, desarrollo de lechones fetales y partos
El estrés oxidativo representa uno de los principales puntos críticos que deben enfrentar los sistemas de producción porcina durante las fases de gestación (temprana, media y tardía) y lactancia, en las que se produce daño sistémico en respuesta al alto nivel de especies reactivas del oxígeno (ERO)(41,50). La gestación refleja un estado de alto estrés oxidativo en el que diversos tejidos y órganos que metabolizan activamente el oxígeno (O2), como la placenta, pueden presentar cambios dinámicos que modifican el estado metabólico tanto de la cerda como del feto. Este estado responde a un aumento en el consumo de O2 y en los niveles de energía necesarios para soportar la carga metabólica involucrada en el desarrollo placentario, embrionario y crecimiento fetal, lo que implica la movilización de tejidos. Sin embargo, un exceso de ERO perjudica la función fisiológica de la placenta, desencadenando restricciones uterinas, que aumentan las tasas de mortalidad embrionaria y perjudican el desarrollo de la progenie(50).
Por ello, para mitigar el impacto del estrés oxidativo, se han incluido diferentes fuentes de antioxidantes en las dietas de las cerdas gestantes. En este contexto, estudios previos(50) han determinado que la inclusión de 200 mg/kg y 300 mg/kg de polifenoles de semilla de uva (PSU) en cerdas multíparas durante la gestación tardía (día 80) redujo la muerte fetal (0.63) y aumentó la supervivencia al parto (89.32 %). Estos resultados reflejaron mayores niveles circulantes de progesterona y estrógeno en suero de cerdas gestantes (día 110), que se asocian con el mantenimiento de la gestación y el reconocimiento materno(50). Asimismo, esta suplementación mostró una mejora significativa en el estado antioxidante de las cerdas gestantes al aumentar la actividad enzimática de la superóxido dismutasa (SOD) y la glutatión peroxidasa (GSHpx)(41,50). Del mismo modo, la suplementación de resveratrol (300 mg/kg) y catequinas (200 y 300 mg/kg) desde el inicio (día 20) y media gestación (día 40) hasta el parto mejoró el estado antioxidante (SOD, GSHpx y catalasa [CAT]) de las cerdas y su progenie durante la fase de lactación(51,52). Estos resultados se atribuyeron a las vías Keap1-Nrf2 y Sirt 1 que regulan los genes antioxidantes placentarios(41). Este mismo comportamiento también ha sido reportado con la inclusión de otros extractos vegetales, lo que reafirma la importancia de una adecuada suplementación materna a partir de cualquier tercio de gestación para asegurar un adecuado desempeño en el parto y durante la lactancia(53).
Fase de lactación: cerdas y lechones lactantes
Al igual que el final de la gestación y el parto, la lactancia también conduce a una sobreproducción de ERO, lo que desencadena la peroxidación lipídica. En este caso, el estrés oxidativo se atribuye a la mayor demanda de energía metabólica necesaria para el desarrollo de la glándula mamaria y la producción de leche. Además, la gravedad del daño oxidativo afecta al desempeño reproductivo de la cerda y dificulta el desarrollo adecuado de la descendencia durante las primeras etapas de la vida. Por lo tanto, la suplementación materna con compuestos fenólicos desde la gestación hasta la lactancia ha sido evaluada como una estrategia para mejorar el estado antioxidante en cerdas lactantes, lo que a su vez puede conducir a un mejor desarrollo, desempeño pre-destete y salud general de su progenie (lechones lactantes)(53).
En este contexto, se ha determinado que la inclusión de PSU (200 y 300 mg/kg), resveratrol (200 y 300 mg/kg) y mezcla estandarizada de polifenoles-Proviox (catequina, procianidinas y antocianinas) aumenta el contenido de inmunoglobulinas (IgG e IgM) en el calostro de las cerdas suplementadas(50), mejorando así el estado antioxidante de los lechones lactantes al potenciar la actividad enzimática de la CAT, SOD y GPx(41,54,55). Además, la tasa de supervivencia pre-destete mejoró (96.9 %) con la suplementación con PSU (200 y 300 mg/kg), mientras que la inclusión de resveratrol en las dietas de las cerdas aumentó el peso de la camada (57.26 kg frente a 48.98 kg), al igual que el peso de los lechones al destete (5.84 kg frente a 5.24 kg), en comparación con las hembras no suplementadas(41,50). De igual forma, con la inclusión de Proviox, se observó un efecto significativo en el peso de la camada de lechones lactantes y el calostro(54). Por lo tanto, las estrategias nutricionales que se han evaluado en esta etapa están enfocadas a mejorar el estado antioxidante de las cerdas gestantes, y así atenuar los impactos del estrés oxidativo, lo cual puede estar asociado a una mejora en el estado inmunológico y esta información sugiere que es posible obtener un mayor número de lechones para las fases de crecimiento y finalización.
Fases de destete y posdestete
La etapa de destete representa un cambio en la dieta de los lechones (de líquida a sólida) y la separación de la madre. Además, el establecimiento de jerarquías sociales desencadena una serie de respuestas fisiológicas que comprometen la función inmune, el metabolismo intestinal y la capacidad antioxidante, favoreciendo la aparición de estrés oxidativo y un aumento en las tasas de morbilidad y mortalidad(56). Al mismo tiempo, la ingesta de alimento disminuye, lo que provoca un retraso en el crecimiento. La adición de SPV se ha evaluado como un factor atenuante para esta situación y como una alternativa para reemplazar el uso de antibióticos como promotores de crecimiento en esta fase(19). En este contexto, se ha demostrado que la adición de CF como el ácido tánico (500 mg/kg y 1000 mg/kg) en las dietas de lechones destetados (3 semanas de edad durante 14 días) tiene efectos beneficiosos sobre el transporte de nutrientes a nivel intestinal junto con la microbiota y morfología intestinal(19,57). Esto refleja el potencial de OU como fuente de varios CF. Un estudio previo determinó que la inclusión de un 5 % de OU seco mejoró el estado antioxidante del hígado, el riñón y el bazo en lechones destetados. Se observó reducción de la peroxidación lipídica (MDA) y de triglicéridos en el día 36, mientras que las actividades de CAT, GSHpx y SOD aumentaron(58). Otros autores han reportado efectos similares sobre la actividad enzimática y la capacidad antioxidante total (CAO-T), como se presenta en el Cuadro 2.
Un estudio reciente(59) reportó que la inclusión de harina de semilla de uva (8 %) en las dietas de inicio atenúa el estrés oxidativo inducido por la aflatoxina B1 (AFB1) y disminuye los marcadores de inflamación (Cuadro 2). En este contexto, se ha demostrado un efecto protector contra la aflatoxicosis y el estado antioxidante de las aves mejoró con la suplementación con resveratrol (0.5 y 1 %)(58). Otros estudios han reportado que los subproductos agroindustriales como el OU blanco (Malvasía) y el OU rojo (Primitivo) tienen una excelente capacidad para adsorber ocratoxina (OTA), zearalenona (ZEN), fumonisina (FB1) y AFB1 de soluciones acuosas a pH 3 y 7(60). Estos resultados muestran que estos compuestos tienen un potencial considerable para secuestrar micotoxinas en dietas de lechones destetados. El estudio demostró que la inclusión de orujo blanco (Malvasía 2.8 %) en la dieta de cerdos destetados reduce la absorción gastrointestinal de las micotoxinas ZEN y OTA en un 67 y 69 %, respectivamente. Aunque aún no se ha dilucidado el mecanismo de acción como secuestrante de micotoxinas, se sugiere que el contenido de celulosa en el orujo tiene un potencial considerable para absorber AFB1 a través de atracciones electrostáticas; los polifenoles también pueden formar complejos con micotoxinas(61).
Otras actividades biológicas, como la inmunomoduladora, la antiinflamatoria y la modulación intestinal, se han evaluado con mayor frecuencia en lechones destetados, dados sus órganos digestivos y sistemas inmunes inmaduros. Se ha enfatizado la evaluación en las primeras 3 semanas después del destete, ya que el sistema inmune solo puede generar una respuesta efectiva a las 5 o 6 semanas de edad(62,63). En este contexto, se ha demostrado que la suplementación con 50, 100 y 150 mg/kg de procianidinas de semilla de uva(46); 1 % de extracto de semilla de uva y orujo de uva (ESUOU) con un contenido de fenoles del 8.5 % (lechones de 7 kg de peso corporal durante 4 semanas)(49,64); y 5 % de OU seco (20.41 mg/g de materia seca, 5 semanas de edad durante 28 días) disminuye la incidencia de diarrea(58,62), aumenta la relación altura de las vellosidades intestinales: profundidad de la cripta en el duodeno (RVC)(62) y yeyuno(65); aumenta la proporción de Olsenella umbonata, Lactobacillus delbrueckii y Selenomonas bovis en el ciego(65); disminuye las poblaciones de Streptococcus y Clostridium; y disminuye los niveles de AGV en la microbiota fecal de lechones destetados (5 semanas de edad durante 25 días)(58).
Estos resultados se atribuyeron al potencial de los CF para mejorar el estado antioxidante, reducir la permeabilidad intestinal, aumentar el área superficial, mejorar la absorción de nutrientes y modular las poblaciones intestinales mediante el aumento de especies productoras de butirato que mejoran la salud del colon. Estos efectos benéficos podrían estar relacionados con metabolitos microbianos como el ácido 4-hidroxifenilvalérico, 3-hidroxibenzoico, cafeico, siríngico y protocatecuico(66). Del mismo modo, otros estudios han verificado que los CF de SPV reducen la expresión de genes proinflamatorios en el intestino (ciego, íleon y colon)(67). La suplementación con HSU (harina de semilla de uva) en lechones destetados en un modelo de colitis inducida (sulfato de hierro dextrano) atenuó el efecto de este inductor, moduló la microbiota del colon y redujo el impacto de la disbiosis intestinal(25,27). Por lo tanto, los CF de los SPV representan una estrategia para inhibir la inflamación y modular la salud intestinal durante la etapa posdestete. Entre los parámetros productivos, se observó una mejora significativa de 4 a 7 % en la relación ganancia de peso:alimento (G:A)(67), y se ha sugerido que esto se debe a una mejora en la digestibilidad de los nutrientes(25). Sin embargo, otros autores no obtuvieron una mejoría significativa en este indicador y afirmaron que no hubo un efecto consistente sobre la morfología intestinal(68). Por lo tanto, la mejora en G:A se ha asociado con una reducción de la expresión de genes proinflamatorios y cambios en la composición microbiana.
Etapas de crecimiento y finalización
Estudios previos han demostrado que las tortas de orujo de semilla de uva (TSU 5 % de la dieta basal), como fuente rica en proantocianidinas, estilbenos y flavanoles (catequinas, epicatequinas, galocatequinas, epigalocatequinas y procianidinas), aumentaron los niveles de IgA (49.9 %) en el plasma animal(41). Además, el orujo disminuyó la respuesta inflamatoria en el hígado y el bazo, junto con la producción de citoquinas hepáticas (IFN-γ, IL-1, IL-8 e IL-6), y la expresión génica y la concentración de marcadores proinflamatorios (IL-1β e IFN-γ) en el bazo(63). De manera similar, con la misma dosis, se reportó que los niveles de colesterol se redujeron, lo que se atribuye a la capacidad del resveratrol y la epigalocatequina para unirse a reguladores clave del metabolismo lipídico hepático(69). Aunque los niveles de malondialdehído (MDA) disminuyeron en el hígado (13 %), los resultados fueron inconsistentes en términos de expresión de genes enzimáticos; actividades CAT, SOD y GPx, CAO-T; lo que indica que el estado antioxidante de los cerdos no mejoró durante la suplementación y puede atribuirse al bajo contenido de CF en la matriz. En contraste, otros autores(63,68) reportaron un aumento significativo en la expresión de genes enzimáticos (CAT, GPx); una tendencia similar en la actividad de CAT, SOD, GPx y CAO-T (49 %); y disminución de los niveles de MDA en el bazo (20 %). Del mismo modo, con la adición de 9 % de ensilado de OU durante la etapa de crecimiento, la descomposición de H2O2 aumentó(20).
Se ha indicado que la inclusión de 1 % de ESUOU con un contenido de polifenoles de 8.5 % y la adición de 3 % de OU fermentado es eficaz para mejorar el rendimiento productivo (peso corporal, ganancia media diaria [GMD], ingesta de alimento y conversión alimenticia) en cerdos en crecimiento(70). Sin embargo, también se ha demostrado que la inclusión de TSU (5 %) en cerdos en finalización no influye en el desempeño productivo(64). Resultados similares fueron obtenidos por otros autores en todas las etapas de finalización(62). A pesar de que se ha reportado que el OU mejora las capacidades sensoriales y el metabolismo en cerdos(58), pocos estudios específicos para la etapa de finalización (> 70 kg) han evaluado algún subproducto de la vinificación, a diferencia de las dos primeras fases de producción(71). En general, las investigaciones integran la fase de iniciación con el crecimiento(27). Incluso los resultados de la suplementación con OU sobre el desempeño productivo (GMD, conversión alimenticia y consumo de alimento) son inconsistentes(17) y se ha sugerido que esta matriz nutricional puede suprimir el rendimiento del crecimiento. La concentración y el perfil de los CF determinarán este impacto, junto con la unión a las enzimas digestivas y a las proteínas(72).
Un estudio en cerdos reportó que la inclusión de 300 y 600 mg/kg de resveratrol en las dietas de finalización durante 49 d indujo una transición en el tipo de fibras musculares y un cambio en el metabolismo energético(73) que favoreció la expresión de fibras oxidativas (IIA) sobre fibras glucolíticas (IIB). Estos resultados son consistentes con los reportados por otros autores, quienes observaron una menor proporción de fibras IIB cuando los animales fueron suplementados con 400 ppm y 600 ppm de resveratrol durante 42 d, lo que podría indicar un efecto promotor del crecimiento(74). No obstante, los autores no observaron un efecto significativo en la eficiencia de la producción. Asimismo, se ha demostrado que con la inclusión de orujo seco (5 %) en cerdos en finalización, se modifica el perfil de polifenoles en plasma (273 nm y 279 nm) y estos cambios podrían reflejarse en el músculo(75).
Efectos de los subproductos de la vinificación y los extractos de polifenoles del vino en la calidad de la carne de cerdo, el valor nutricional y los productos cárnicos
La suplementación con orujo de uva en dietas o su inclusión directa en carnes o productos cárnicos ejerce un efecto antioxidante y antimicrobiano que determina la calidad final del producto. Asimismo, se ha reportado que la estabilidad oxidativa de los productos cárnicos está determinada por la composición de ácidos grasos insaturados(76,77). A pesar de que un mayor contenido de AGPI en la carne se ha asociado con una mayor susceptibilidad a la oxidación lipídica, también se ha demostrado que la inclusión de fuentes ricas en CF le confiere mayor estabilidad, lo que se atribuye a un aumento de la actividad enzimática antioxidante y a una disminución de MDA y TBARS en el músculo de monogástricos(1,3). Los efectos antioxidantes de los subproductos del orujo, como las semillas y las pieles de uva, que se han visto en la producción animal, se han asociado con productos finales de la degradación de CF de bajo peso molecular, que pueden ser transferidos a los tejidos y se han detectado compuestos responsables como la epicatequina, entre otros(51,76).
Por lo tanto, ha recibido una atención significativa como conservante natural durante el almacenamiento de la carne para prolongar la vida de anaquel. De hecho, se ha demostrado que la suplementación oral de flavonoides (quercetina, epicatequina y catequina) en la dieta aumenta las concentraciones in vivo de vitamina E(78), lo que podría explicar el efecto antioxidante de los CF en los tejidos animales. Además, la dinámica de almacenamiento y deposición de vitamina E difiere en los animales monogástricos. En cerdos, la acumulación de esta vitamina en respuesta a la suplementación con orujo de uva es significativa en grasa, hígado y músculo, mientras que en aves hay más deposición en el hígado y menor deposición en la grasa(79). Además, se ha reportado que la inclusión de 30 g de orujo fermentado/kg en las dietas de cerdos mejoró el porcentaje de marmoleado (3.5 %), aumentó los valores de a* (25 %) y b* (45 %) de la carne, y redujo los niveles de MDA en un 53 %(27). Del mismo modo, el orujo modifica el contenido de ácidos grasos poliinsaturados (AGPI) y la relación AGS/AGPI, lo que se ha atribuido al efecto moderador del orujo de uva sobre la vitamina E en el intestino(78). Asimismo, también se ha demostrado que el suministro de 5 % de orujo de uva de la alimentación de cerdos en finalización aumenta los niveles de ácidos grasos omega 3 (1.27 veces), alfa-linoleico (1.35 veces) y ácido eicosapentaenoico (1.30 veces), mientras que tiende a disminuir la relación n-6:n-3 con respecto al testigo(75).
Además, se ha demostrado que el valor de b* se ve afectado principalmente por el tipo de mioglobina en el músculo(27). Sin embargo, la suplementación de 3 %, 5 %, 6 % y 10 % de orujo de uva con una mezcla de aceites desde la etapa de finalización no afectó la producción de sustancias reactivas al ácido tiobarbitúrico (TBARS, por sus siglas en inglés) en las muestras de lomo de cerdo(79), pero aumentó el valor de a* junto con el índice de saturación de color, lo que sugiere un posible efecto antioxidante.
En cuanto a la textura de la carne, se han obtenido resultados satisfactorios en la fuerza de cizallamiento, que se han asociado a un aumento de los valores de pH en respuesta a la suplementación con extractos de orujo de uva. Además, se ha propuesto que la proantocianidina de uva puede disminuir la deposición de colágeno al suprimir la expresión de promotores proteicos involucrados en la síntesis de colágeno (TGF-β)(80).
Por otro lado, en la maduración de los productos cárnicos, se ha determinado que por cada 100 g de carne cruda se debe utilizar una mezcla de orujo de uva en polvo (1L) a 4 °C durante 72 h. Marinando el lomo de cerdo con esta mezcla, la proteína cruda, la grasa cruda, las cenizas, el pH, el valor de a* y el valor de b* en la carne disminuyeron, y su fuerza de cizallamiento y contenido de humedad aumentaron. Durante el almacenamiento de la carne, la concentración de TBARS, nitrógeno básico volátil y el recuento microbiano total disminuyeron(80,81). Estos resultados podrían ser atribuidos a los ácidos orgánicos y CF presentes en los SPV, que podrían inhibir el crecimiento de microorganismos al reducir la actividad de algunas enzimas e inhibir la producción de sustancias básicas(82). Por lo tanto, marinar con orujo de uva en polvo mejora algunas características de calidad de la carne y estabiliza el lomo de cerdo durante el almacenamiento. Resultados similares se observaron al agregar dos diferentes extractos de orujo de uva a hamburguesas de cerdo(83), lo que resultó en una inhibición en la oxidación lipídica, una mayor estabilidad del color y una mejor aceptabilidad con el método de extracción HLIP (Cuadro 3).
Cuadro 3: Adición de subproductos de vinificación en la carne de cerdo y productos porcinos
* Suplementación de piensos para animales.
Consideraciones tecnológicas para la inclusión de subproductos de la vinificación en dietas de monogástricos
Los SPV como el orujo de uva han sido sometidos a diversos pretratamientos (enzimas endógenas, enzimas celulolíticas, polietilenglicol y fermentación) para aumentar la biodisponibilidad de sus CB, especialmente de los CF, y mejorar su valor nutricional para favorecer su inclusión en la alimentación animal. En este contexto, se ha demostrado que la inclusión de complejos enzimáticos (tanasas) y polietilenglicol en dietas de monogástricos puede inactivar parcialmente los taninos condensados del orujo de uva, minimizar el contenido de factores antinutricionales y aumentar la actividad antioxidante y antimicrobiana(28,84). Entre estos tratamientos, se han destacado los procesos de fermentación con levaduras, bacterias y hongos(27). En este contexto, se desarrolló un enfoque biotecnológico sistemático, a través del cual se evaluaron las capacidades metabólicas de varias especies de hongos (Aspergillus, Rhizopus y Trichoderma ssp.) sobre la bioconversión de orujo de uva y lías a alimento para animales(85). En este estudio, los autores obtuvieron una mejora en el contenido de proteínas (5 a 26 %) y en la digestibilidad del alimento (25 a 50 %).
Del mismo modo, la suplementación de 3 % de orujo de uva fermentado con Saccharomyces boulardii en cerdos durante su etapa de crecimiento-finalización aumentó la digestibilidad aparente de la materia seca (79 %) y el nitrógeno (82.5 %) después de 11 semanas(27). Estos resultados sugieren que los CF de esta matriz pueden estimular la fermentación intestinal e influir en la producción de metabolitos microbianos específicos, lo que explica la mejora en el crecimiento. Esta estrategia posibilita el suministro de fuentes ricas en proteínas y un estado nutricional óptimo a los animales(66). No obstante, también se ha reportado que con la inclusión de 20 % de orujo de uva fermentado en cerdos en finalización, se redujo la digestibilidad de la materia seca, proteína cruda, energía y aminoácidos esenciales y no esenciales con respecto al testigo(25). Estos resultados se atribuyeron a la formación de complejos entre algunas CF y la proteína del alimento, lo que limitó la acción de las enzimas digestivas; por lo tanto, es necesario considerar los niveles de inclusión para cada etapa de producción.
Otros procesos tecnológicos como la elaboración de harinas, polvos y tortas a partir de SPV (semillas, orujo, pieles) son indispensables en la formulación de dietas para animales monogástricos. Por ello, se han evaluado y estandarizado las condiciones óptimas de temperatura, tiempo y tamaño de partícula para obtener mejores resultados en la alimentación animal con respecto al subproducto crudo. El OU ha sido sometido a un proceso de secado con flujo de aire caliente a 65 °C durante 4 días, 60 °C durante 20 h(16,86), 50 °C durante 12 h(87) y 60 °C hasta alcanzar un peso constante. Además, después de la molienda, se han definido tamaños de partícula de 1 mm a 6 mm para aves y cerdos respectivamente(67,70,86). Asimismo, también es importante definir y especificar las condiciones de temperatura/tiempo para el OU destinado a la alimentación de cerdos y estandarizar los tamaños de partícula dependiendo de la etapa de producción, lo que ha permitido obtener mejores resultados en la alimentación animal con respecto a los subproductos crudos(8,20,70).
Conclusiones e implicaciones
Un gran número de estudios realizados han evaluado las propiedades antioxidantes, antiinflamatorias, antimicrobianas e inmunomoduladoras de los subproductos de la industria vitivinícola, con énfasis en lechones destetados y primeras etapas de crecimiento. Sin embargo, no se ha determinado el efecto sobre las fibras musculares con la matriz cruda. En la etapa final de la producción, aún es necesario continuar evaluando diferentes presentaciones de esta matriz y nuevos niveles de inclusión y considerar las tasas de mortalidad como otra medida indirecta del desempeño productivo. La inclusión de orujo de uva en forma de extracto o harina y sus CB en las dietas de los cerdos en cada fase productiva se considera una fuente antioxidante y antimicrobiana de bajo costo que ejerce múltiples beneficios sobre el desempeño productivo de los cerdos, así como sobre la calidad de la carne. Por lo tanto, representa una alternativa prometedora para el sector de nutrición animal que podría minimizar el uso de compuestos antioxidantes sintéticos, promotores de crecimiento como antibióticos y agentes beta-adrenérgicos en el sistema de producción porcina.
Conflicto de intereses
Los autores declaran que no existen conflictos de intereses.
Literatura citada:
Cuadro 2: Efectos de los compuestos fenólicos de los subproductos de la vinificación en los sistemas de producción porcina
a) Tipo de aditivo: Z= zootécnico; T= tecnológico; S= sensorial; DP= desempeño productivo; DR= desempeño reproductivo; CAO-T (capacidad antioxidante total).
b) Resultados: MDA (malondialdehído); EAO (estado antioxidante); NR= no reportado, MFI= modulación de la función intestinal; CDA= coeficiente de digestibilidad aparente.