https://doi.org/10.22319/rmcp.v15i3.6456
Artícle
Corn oil in Pelibuey ewes embryo transfer
Christofer Israel Márquez Hernández a
Arturo Pro Martínez b
Glafiro Torres Hernández b
Raymundo Rangel Santos a
Jaime Gallegos Sánchez b *
a Universidad Autónoma Chapingo. México.
b Colegio de Posgraduados. Programa de Ganadería, Montecillo, Texcoco, Estado de México. México.
* Corresponding author: gallegos@colpos.mx
Abstract:
The effect of including corn oil (CO) in the diet of Pelibuey ewes in an embryo transfer protocol was evaluated. Twenty-four (24) donor ewes were randomly assigned to each of two groups (G), G1: twelve ewes fed a base diet (BD) with CO, and G2: twelve ewes fed BD without CO. Embryos were obtained 6.5 d after estrus and transferred to recipient ewes, forming four treatments: T1: recipient ewes fed a base diet with CO (BD+CO), transferred with embryos from a G1 donor (n=23), T2: recipient ewes fed BD without CO (BD-CO), transferred with embryos from a G1 donor (n=18), T3: recipient ewes fed BD+CO, transferred with embryos from a G2 donor (n=9), and T4: recipient ewes fed BD-CO, transferred with embryos from a G2 donor (n=11). The inclusion of CO in the diet increased (P<0.05) the ovulatory rate (10.5 ± 2.07 vs. 6.3 ± 2.07 corpora lutea), the number of transferable embryos (5.5 ± 1.4 vs 2.8 ± 1.4), quality 1 embryos (4.41 ± 1.1 vs 2.08 ± 1.1), and the number of total structures (5.9 ± 1.5 vs 3.1 ± 2.7) per donor ewe, and no differences (P>0.05) were found in the percentage of gestation (43.4, 55.5, 55.5, and 36.3 %) in the recipient ewes for any of the treatments. Including CO in the diet of Pelibuey ewes increases the superovulatory response in an embryo transfer protocol.
Keywords: Corn oil, Pelibuey ewes, Embryos.
Received: 02/05/2023
Accepted: 15/05/2024
Introduction
Reproductive technologies are valuable tools that help increase sheep production units’ productive and economic efficiency. For example, embryo transfer (ET) has been successfully used to reproduce genetically superior animals(1). Nevertheless, even though this technology has advanced considerably in recent decades, the response to superstimulation protocols and the percentage of fertility have been inconsistent, so research has focused on two fundamental aspects: Improving the response in follicular superstimulation of embryo donor females to obtain a bigger number of transferable embryos and increasing the percentage of gestation in recipient females.
Some specific diet components, such as energy, can positively influence certain reproductive aspects of the female(2). Supplementation with corn oil (CO) with a high content of polyunsaturated fatty acids (PUFAs; oleic, linoleic, and linolenic) can be used to modify some processes of ovarian physiology in ewes(3) because these processes are related to energy availability(4). It has been shown that dietary CO supplementation can improve the population of large follicles(5) and promote the number of corpora lutea(6) and prolificacy(7). PUFAs have a more substantial effect in the early stages of folliculogenesis and, therefore, can increase the number of embryos obtained by donor females and promote the conception rate in dairy cows(8). Based on this background, this research aimed to determine if the inclusion of CO in the diet of Pelibuey ewes improves the superstimulatory response, increases the number of transferable embryos in donor ewes and improves the pregnancy rate of recipient ewes in an ET program in Pelibuey ewes.
Material and methods
The study was carried out from July to December 2017 at the Sheep and Goat Reproduction Laboratory (LaROCa, for its acronym in Spanish) of the College of Postgraduates, Montecillo Campus, located at 19° 29’ N, 98° 53’ W, and 2,240 m asl(9).
Twenty-four (24) Pelibuey donor ewes aged 3.5 ± 0.3 yr with an average weight of 51.9 ± 3.2 kg were used. Of this group, 12 ewes received a base diet with corn oil (G1) at a rate of 2.0 kg ewe-1 d-1 consisting of 60 % oat hay, 25 % achicalada (sun-dried) alfalfa, 6 % corn oil (Mazola®), and 9 % commercial concentrate (Borrega plus: Alimentos Unión Tepexpan®); the rest of the ewes (12) received the following diet without corn oil (G2): 63 % oat hay and 26 % achicalada alfalfa, and 11 % commercial concentrate (Borrega plus: Alimentos Unión Tepexpan®; 12 % CP, 40 % ADF, 51 % NDF, 2.3 % EE, and 9 % ash); the experimental diets were isoenergetic and isoproteic, containing 12 % CP and 3.2 Mcal ME, and were offered for 16 d from day three of the estrus synchronization protocol until the day before transfer (the time of CIDR insertion was considered day 0).
As recipient ewes, 61 Pelibuey ewes aged 3.4 ± 0.3 yr with an average weight of 54.6 ± 1.2 kg were used. Thirty-two (32) ewes received the same diet as G1 at a rate of 2.0 kg ewe-1 d-1; the rest (29) of the ewes received the same diet as G2. The experimental diets were offered for 16 d from day three of the estrus synchronization protocol until the day before the transfer (the time of CIDR insertion was considered as day 0). The treatments were assigned as follows (Table 1):
Table 1: Allocation of experimental treatments in recipients
*BD+CO= Base diet with corn oil, BD-CO=Base diet without corn oil.
The number of recipient females in the treatments was subject to the availability of the transferable embryos at the time of donor flushing.
All the ewes were handled according to the standards established by the “Regulation for the use and care of animals intended for research in the College of Postgraduates” and the Official Mexican Standard NOM-062-ZOO-1999 in order to avoid unnecessary stress and suffering of the animals.
All recipient ewes were subjected to an estrus synchronization protocol by insertion of an intravaginal device (CIDR-Pfizer®) impregnated with progesterone (0.3 g of P4) for 9 d; on day seven, 300 IU of equine chorionic gonadotropin (eCG) (Folligon-Intervet ®) and a dose of 5 mg of prostaglandin F2α (Lutalyse®, Pharmacia Animal Health Laboratories) were applied via IM. The timing in the donors was similar, only the IM application of 200 mg in decreasing doses (40-40, 30-30, 20-20, 10-10 mg) of follicle-stimulating hormone (FOLLTROPIN®, Vetoquinol Laboratories) at 12-h intervals between each application (am-pm) was included on d 6 to 9 of CIDR insertion for the superstimulation process.
On d 9 of the synchronization protocol, the CIDR was removed, and the detection of females in estrus began 4 h after its removal with the help of a ram provided with an apron in order to avoid copulation. Donor ewes that presented estrus behavior were inseminated intrauterinely (laparoscopy) from 12 to 18 h after the onset of estrus and remained without feed for 24 h, a procedure to reduce rumen content and avoid bronchoaspiration(10). For insemination, two straws of 0.25 ml of fresh semen containing 80 x 106 sperm per straw were used.
Embryo collection and transfer
Ewes that presented a superstimulatory response (>2 corpora lutea(11)) underwent embryo collection 6.5 d after estrus by means of ventral midline laparotomy(12). To do this, the ewes were exposed to a 24-h fast before the procedure, and the number of CL present in each ovary was recorded. An anesthesia protocol with a combination of xylazine (Procin®, Pisa Laboratories) and ketamine (Anesket®, Pisa Laboratories) in a 0.8:0.2 dilution was applied intravenously in the jugular vein. Subsequently, the ewe was prepared on the stretcher placed on a plane at 45°, and 1 ml of lidocaine (Pisacaina®, Pisa Laboratories) was applied. The surgical drapes were placed, a 7 cm incision was made on the ventral midline, and the reproductive system was exposed with Babcock forceps. With the help of an intravenous catheter (Punzocat®, 32 mm), a puncture was made in the uterus-tubal junction to enter the lumen of the uterine horn; a second puncture was then performed at the level of the intercornual ligament to introduce a Foley No. 10 catheter. A syringe with 60 ml of a flushing medium (Vigro® Complete Flush Solution, Bioniche Laboratories) at 37 °C was connected to the intravenous catheter so that the embryos were collected by dragging them into gridded Petri dishes with the solution that came out of the front end of the Foley catheter. The Petri dishes were transferred to the laboratory to begin the search and evaluation of the embryos.|
The embryo search was carried out meticulously with a stereoscopic microscope (Barnstead, USA) at 40X magnification, observing each quadrant of the Petri dish. The structures found were placed in a four-well Petri dish containing 200 μL of holding medium (Holding®, Bioniche Laboratories) in each well, placed in a thermal stage at 37 °C. The embryos were evaluated and classified under the stereoscopic microscope at 100X magnification according to their morphology and based on the criteria of the International Embryo Transfer Society (IETS)(13).
The embryos were transferred fresh six and a half days after the onset of estrus (trying to reduce the asynchrony time between the donor and the recipient), selecting females from both the BD+CO group and the BD-CO group, making sure that all the females in the four treatments had the same number and quality of embryos transferred. With a previous period of 24 h of fasting, the recipient was placed on a stretcher, and the same protocol as for embryo collection was carried out. The uterine horns were visualized laparoscopically, and by means of a ventral midline laparotomy, a 2 cm incision was made in the abdominal midline, and with a Babcock forceps, the ipsilateral uterine horn of the ovary with one or more normal corpora lutea was externalized, permanently hydrating it with physiological saline solution (CS® Solution, Pisa Laboratories). With the help of an intravenous catheter (Punzocat®, 32 mm), a puncture was made in the uterus-tubal junction, and an embryo of quality 1 or 2 (depending on availability) was placed in a catheter (TOMCAT®) under the following order: a column of medium, air, medium with two embryos, air and finally medium. The embryos placed in the catheter were propelled into the lumen of the uterine horn using an insulin syringe. The surgical planes were sutured with zero-gauge chromic Catgut® for peritoneum and muscle, and the skin was sutured with zero-gauge nylon. To prevent postoperative infections and once the suture was finished, 100 mg of oxytetracycline (Emicina LA®, Pfizer Laboratories) was applied via IM.
The variables evaluated were the following:
Estrus manifestation (%): percentage of ewes that showed signs of estrus within each treatment.
Time to estrus manifestation (h): hours elapsed from removal of the device to the occurrence of estrus.
Ovulatory rate (OVR): number of corpora lutea present at the time of embryo extraction (6.5 days after estrus).
Quality 1 embryos (EQ1): number of quality 1 morulae or blastocysts.
Quality 2 embryos (EQ2): number of quality 2 morulae or blastocysts.
Transferable embryos (TE): number of quality 1 and 2 morulae or blastocysts.
Oocytes (OO): number of oocytes that were not fertilized.
Total structures (TS): number of total collected structures.
Percentage of gestation: number of pregnant recipient ewes at 40 d post-transfer divided by the number of ewes in each treatment multiplied by 100.
Lambing rate: number of recipient ewes that lambed in relation to the number of ewes transferred in each treatment.
The response in donor ewes was analyzed using a Poisson regression using the Generalized Linear Model (PROC GENMOD). To analyze the percentage of pregnancy in the recipients, the analysis of contingency tables for two factors (DAM and RAM) was employed, using the Log-linear model through the CADMOD procedure of the SAS statistical package(14). The manifestation of estrus was analyzed through a Chi-square independence test using the PROC FREC procedure, and the time to estrus manifestation was analyzed with the Kaplan Meier survival curves method using the LIFETEST procedure, and differences were established between treatments with the LOG RANK test.
Results
The proportion of donor ewes with signs of estrus and the time to the estrus were similar between treatments (P>0.05); however, the inclusion of CO in the diet of donor ewes increased (P<0.05) OVR, TE, EQ1, OO, and the number of TS per ewe (Table 2).
Table 2: Response of Pelibuey donor ewes fed a base diet with corn oil (BD+CO) or with a base diet without corn oil (BD-CO) (Means ± SE)
ab Values with different literal within rows show differences (P<0.05).
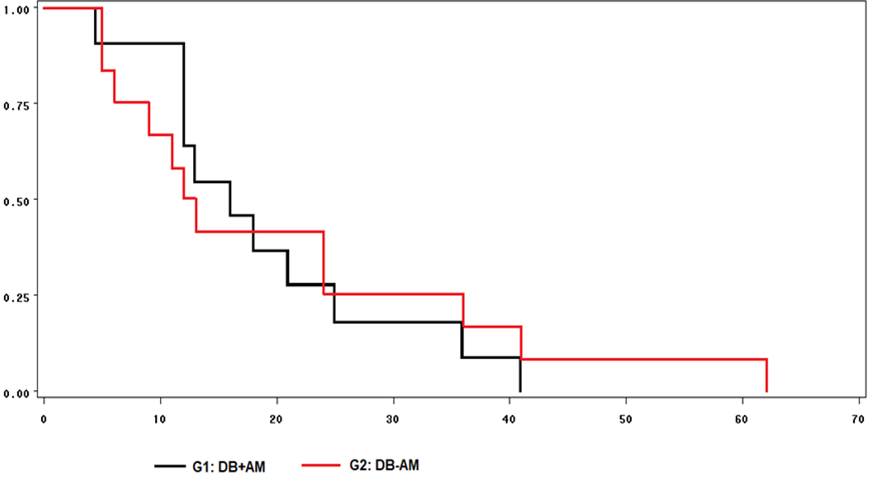
No differences were observed in the probability of estrus manifestation between groups, obtained by means of survival curves (Figure 1). Thus, 100 % of the ewes with BD+CO manifested estrus 42 h after CIDR removal, while the BD-CO ewes did so until 62 h.
Figure 1: Survival curves for the time between CIDR removal and estrus manifestation in Pelibuey donor ewes fed a base diet with corn oil (BD+CO) or with a base diet without corn oil (BD-CO)
Response in recipients
The inclusion of CO in the diet did not alter the time or number of recipient ewes in estrus
with a base diet with corn oil (BD+CO, 24.08 ± 1.03) or with a base diet without corn oil (BD-CO, 25.8±3.0).
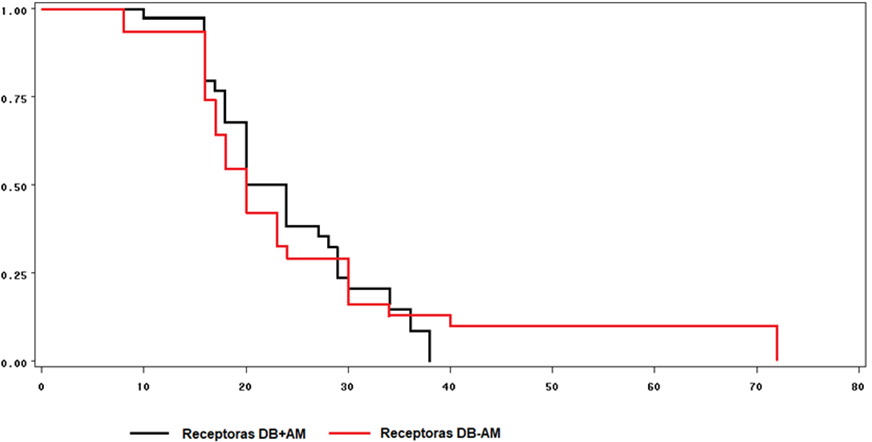
Survival curves for estrus manifestation in recipients showed no differences between treatments (Figure 2). One hundred percent of the ewes with CO manifested estrus at 38 h after CIDR removal, while the control ewes did so at 72 h.
Figure 2: Survival curves for the time between CIDR removal and estrus manifestation in Pelibuey recipient ewes fed a base diet with corn oil (BD+CO) or a base diet without corn oil (BD-CO)
No differences were found (P>0.05) in the percentages of pregnancy and lambing for any of the recipients’ treatments (Table 3).
Table 3: Response in Pelibuey recipient ewes fed a base diet with corn oil (BD+CO) or with a base diet without corn oil (BD-CO)
(P>0.05).
Discussion
The response to manifestation and time to estrus in donor and recipient Pelibuey ewes did not show differences between treatments (P>0.05). Similarly, in a study with Corriedale ewes, 100 % were reported to have superovulated in estrus with eCG and FSH during the breeding season(15). In the present study, 25 % of donor ewes did not respond to the superovulation protocol, which coincides with other studies in which the proportion of females that did not respond to superovulation was between 20(16) and 30 %(17). Supplementing omega-6 (n-6) PUFAs increases energy consumption as they are essential for feeding ruminants in addition to participating in the processes of folliculogenesis, ovulation, and estrous behavior(18). The population of medium and large follicles increased when corn oil was supplemented as a source of PUFAs in ewes, which may result in increased estrogen production by those follicles, exerting an effect similar to eCG(19). In the present study, the inclusion of PUFAs in the diet of ewes did not modify their reproductive behavior, possibly due to the short time of exposure to PUFAs in the diet since it has been mentioned that, in order to observe a response in the reproductive behavior of females, exposure of at least 20 d before the day of mating and continuing during mating is required(20).
The time to the manifestation of estrus between recipients (24.08 vs 25.8 h) and donors (19.13 vs 20.6 h) was similar between treatments (P>0.05). Results from other authors showed an estrus onset of 34.5.0 ± 2.6 h(21) or 32 ± 5.6 h(22) when 500 IU of eCG was applied, differing from what was found in this study. It has been mentioned that the time to the beginning of estrus is determined by the breed and type of FSH used in superovulation protocols(23). The time to the beginning of estrus was 24 h, shorter when ewes were subjected to an intravenous infusion of olive oil compared to soybean oil. It has been mentioned that olive oil stimulates plasma concentrations of prostaglandin F2 alpha metabolite (PGFM) and prostaglandin E2 (PGE2)(24), providing evidence of the possible relationship between fatty acids, prostaglandins, and the onset of estrus(25). In the present research, it was not possible to observe such an effect, probably due to the hormones used for synchronization.
The inclusion of CO in the diet of donor ewes improved the response to superstimulation in G1, results that are similar to those found by Herrera-Camacho(6) when evaluating superovulated Pelibuey ewes supplemented with CO (4 % DM), observing an increase in the number of corpora lutea, embryos collected and morulae; on the other hand, Zeron(26) reported a higher quality in oocytes and an increase in the proportion of PUFAs in plasma, follicular fluid, and cumulus oophorus cells when supplementing ewes with calcium soaps of fish oil. Nutrition during oocyte maturation has important effects on embryo viability in superovulated ewes(27). FAs can directly affect oocyte maturation through the composition of its membrane(28) or indirectly by affecting the concentration of metabolites in follicular fluid, impacting its subsequent development and viability(29). Therefore, it is suggested that the inclusion of CO in this research could have influenced the quality of oocytes and embryos during the early stages of development since more transferable embryos were obtained when PUFAs were added to the diet; in addition, the increase in the number of corpora lutea may have modified the secretion of progesterone as there was a greater availability of cholesterol for the luteal tissue(6), optimizing the conditions for better embryonic development.
No differences in pregnancy percentage were observed between the four treatments (P>0.05). Similar results were found in other studies where, when adding CO to the diet of Pelibuey ewes, they did not observe differences in the percentage of gestation(7,19). In this regard, it has been mentioned that both undernutrition and overfeeding can cause alterations in the intrauterine environment that prevent adequate development of the embryo, affecting its viability(30).
Diets containing high concentrations of PUFAs, such as linoleic acid in corn oil, can regulate prostaglandin synthesis and its subsequent effects on corpus luteum persistence, E2 synthesis(4,8,25), and maternal recognition of pregnancy(31). In addition, PUFAs can increase circulating concentrations of progesterone due to an increase in the availability of cholesterol(32), the main precursor for the synthesis of progesterone (P4) in the corpus luteum(33).
The response to superovulation found in the present study was similar to that reported in other studies, obtaining an increase in the number of transferable embryos in the donors who received the PUFAs; nevertheless, the addition of PUFAs in the diet of recipient ewes did not improve the percentage of gestation, so it is necessary to carry out more studies on the time of exposure of females to the diet with PUFAs and the quantity and quality of PUFAs offered since, in ewes, there are very few published studies that explain the direct effects of polyunsaturated fatty acids on fertility(25).
Conclusions and implications
In conclusion, donor ewes respond well to including corn oil in an embryo transfer program. The use of corn oil in the diet of Pelibuey ewes increases the ovulatory rate, the number of total structures, and the number of transferable embryos, but it has no effect on the pregnancy percentage of the embryo recipient females.
Acknowledgments
To the Sheep and Goat Reproduction Laboratory (LaROCa, for its acronym in Spanish), Montecillo Campus of the College of Postgraduates, and to the Lines of Generation and Application of Knowledge (LGAC, for its acronym in Spanish) Technological Innovation and Food Safety in Livestock for their financial support.
Literature cited: