https://doi.org/10.22319/rmcp.v15i3.6456
Artículo
Aceite de maíz en la transferencia de embriones de ovejas Pelibuey
Christofer Israel Márquez Hernández a
Arturo Pro Martínez b
Glafiro Torres Hernández b
Raymundo Rangel Santos a
Jaime Gallegos Sánchez b *
a Universidad Autónoma Chapingo. México.
b Colegio de Posgraduados. Programa de Ganadería, Montecillo, Texcoco, Estado de México. México.
* Autor de correspondencia: gallegos@colpos.mx
Resumen:
Se evaluó el efecto de la inclusión de aceite de maíz (AM) en la dieta de ovejas Pelibuey en un protocolo de transferencia de embriones. Veinticuatro ovejas donadoras fueron asignadas al azar a cada uno de dos grupos (G), G1: doce ovejas alimentadas con una dieta base (DB) con AM y, G2: doce ovejas alimentadas con DB sin AM. Los embriones fueron obtenidos 6.5 días después del estro y transferidos a ovejas receptoras formando cuatro tratamientos: T1: receptoras alimentadas con dieta base con AM (DB+AM) transferidas con embriones de una donadora del G1 (n=23), T2: receptoras alimentadas con DB sin AM (DB-AM) transferidas con embriones de una donadora del G1 (n=18), T3: receptoras alimentadas con DB+AM transferidas con embriones de una donadora del G2 (n=9) y T4: receptoras alimentadas con DB-AM transferidas con embriones de una donadora del G2 (n=11). La inclusión de AM en la dieta incrementó (P<0.05) la tasa ovulatoria (10.5 ± 2.07 vs 6.3±2.07 cuerpos lúteos), el número de embriones transferibles (5.5 ± 1.4 vs 2.8 ± 1.4), embriones calidad 1 (4.41 ± 1.1 vs 2.08 ± 1.1) y la cantidad de estructuras totales (5.9 ± 1.5 vs 3.1 ± 2.7) por oveja donadora, y no se encontraron diferencias (P>0.05) en el porcentaje de gestación (43.4, 55.5, 55.5 y 36.3 %) en las receptoras para ninguno de los tratamientos. La inclusión de AM en la dieta de ovejas Pelibuey incrementa la respuesta superovulatoria en un protocolo de transferencia de embriones.
Palabras clave: Aceite de maíz, ovejas Pelibuey, Embriones.
Recibido: 02/05/2023
Aceptado: 15/05/2024
Introducción
Las tecnologías reproductivas son herramientas valiosas, que ayudan a incrementar la eficiencia productiva y económica de las unidades de producción de ovinos. Por ejemplo, la transferencia de embriones (TE) ha sido utilizada exitosamente para reproducir animales genéticamente superiores(1). Sin embargo, a pesar de que esta tecnología ha avanzado considerablemente en las últimas décadas, la respuesta a los protocolos de superestimulación y el porcentaje de fertilidad han sido inconsistentes, por lo que las investigaciones se han enfocado en dos aspectos fundamentales: Mejorar la respuesta en la superestimulación folicular de hembras donadoras de embriones, para obtener un mayor número de embriones transferibles, y aumentar el porcentaje de gestación en las hembras receptoras.
Algunos componentes específicos de la dieta, como la energía, pueden influir positivamente en ciertos aspectos reproductivos de la hembra(2). La suplementación con aceite de maíz (AM) con alto contenido de ácidos grasos poliinsaturados (AGP; oleico, linoleico y linolénico) puede ser utilizado para modificar algunos procesos de la fisiología del ovario en ovejas(3) debido a que estos procesos están relacionados con la disponibilidad de energía(4). Se ha demostrado que la suplementación con AM en la dieta puede mejorar la población de folículos grandes(5) favorece el número de cuerpos lúteos(6) y la prolificidad(7). Los AGP tienen un efecto mayor en las primeras etapas de la foliculogénesis y, por lo tanto, pueden aumentar el número de embriones obtenidos por hembras donadoras y son capaces de favorecer la tasa de concepción en vacas lecheras(8). Con base en estos antecedentes, el objetivo de la investigación fue determinar si la inclusión de AM en la dieta de ovejas Pelibuey mejora la respuesta superestimulatoria e incrementa el número de embriones transferibles en las ovejas donadoras, y mejora el porcentaje de gestación de ovejas receptoras en un programa de TE en ovejas de raza Pelibuey.
Material y métodos
El estudio se realizó durante los meses de julio a diciembre de 2017 en el Laboratorio de Reproducción de Ovinos y Caprinos (LaROCa) del Colegio de Postgraduados, Campus Montecillo, ubicado a 19° 29’ N y 98° 53’ O y 2,240 msnm(9).
Se utilizaron 24 ovejas donadoras Pelibuey de 3.5 ± 0.3 años de edad y un peso promedio de 51.9 ± 3.2 kg. De este grupo, doce de ovejas recibieron una dieta base con aceite de maíz (G1), a razón de 2.0 kg oveja-1 dia-1 que consistió de: 60% heno de avena, 25% alfalfa achicalada, 6% de aceite de maíz (Mazola®) y 9% concentrado comercial (Borrega plus: Alimentos Unión Tepexpan ®); el resto de las ovejas (12) recibieron la siguiente dieta sin aceite de maíz (G2): 63% heno de avena y 26% alfalfa achicalada, 11% concentrado comercial (Borrega plus: Alimentos Unión Tepexpan ®;12 % PC, 40 % FDA, 51 % FDN, 2.3 % EE y 9 % cenizas), las dietas experimentales fueron isoenergéticas e isoprotéicas, contenían 12 % PC y 3.2 Mcal EM, y se ofrecieron durante 16 días, desde el día tres del protocolo de sincronización de estro, hasta el día previo a la transferencia (el momento de la inserción del dispositivo intravaginal CIDR fue considerado día 0).
Como ovejas receptoras se utilizaron 61 ovejas Pelibuey de 3.4 ± 0.3 años de edad y un peso promedio de 54.6 ± 1.2 kg. Treinta y dos (32) ovejas recibieron la misma dieta que G1 a razón de 2.0 kg oveja-1 dia-1, el resto (29) de las ovejas recibieron la alimentación igual que G2. Las dietas experimentales se ofrecieron durante 16 días, desde el día tres del protocolo de sincronización de estro, hasta el día previo a la transferencia (momento de la inserción del CIDR fue considerado como día 0). Los tratamientos quedaron asignados de la siguiente manera (Cuadro 1):
Cuadro 1: Asignación de tratamientos experimentales en receptoras
*DB+AM= Dieta base con aceite de maíz, DB-AM=Dieta base sin aceite de maíz.
El número de hembras receptoras en los tratamientos estuvo sujeto a la disposición de los embriones transferibles al momento del lavado de las donadoras.
Todas las ovejas se manejaron según las normas establecidas por el “Reglamento para el uso y cuidado de animales destinados a la investigación en el Colegio de Postgraduados” y la Norma Oficial Mexicana NOM-062-ZOO-1999, con la finalidad de evitar estrés y el sufrimiento innecesario de los animales.
Todas las ovejas receptoras fueron sometidas a un protocolo de sincronización de estro, mediante la inserción de un dispositivo intravaginal (CIDR®-Pfizer), impregnado con progesterona (0.3 g de P4) durante nueve días, en el día siete se aplicaron por vía IM 300 UI de gonadotropina coriónica equina eCG (Folligon-Intervet ®) además de una dosis vía IM de 5 mg de prostaglandina F2α (Lutalyse®, Laboratorios Pharmacia Animal Health). La sincronización en las donadoras fue similar, solo se incluyó en los días 6 al 9 de la inserción del CIDR, la aplicación im de 200 mg en dosis decrecientes (40-40, 30-30, 20-20, 10-10 mg) de hormona folículo estimulante (FOLLTROPIN® Laboratorios Vetoquinol) en intervalos de 12 h entre cada aplicación (am-pm) para el proceso de superestimulación.
El día nueve del protocolo de sincronización, se retiró el CIDR y la detección de hembras en estro inició 4 h después de su retiro, con el auxilio de un carnero provisto con un mandil con la finalidad de evitar la cópula. Las ovejas donadoras que presentaron conducta de estro, se inseminaron de manera intrauterina (laparoscopía) de 12 a 18 h después del inicio del estro, y permanecieron sin alimento durante 24 h, procedimiento para reducir el contenido ruminal y evitar broncoaspiraciones(10). Para la inseminación se utilizaron dos pajillas de 0.25 ml de semen fresco conteniendo 80 x 106 de espermatozoides por pajilla.
Colección y transferencia de embriones
Las ovejas que presentaron respuesta superestimulatoria (>2 cuerpos lúteos(11)) se sometieron a una colecta de embriones, 6.5 días después del estro, mediante laparotomía media ventral(12). Para ello, las ovejas fueron expuestas a un ayuno previo de 24 h al procedimiento y se registró el número de CL presentes en cada ovario. Se aplicó un protocolo de anestesia con una combinación de xilacina (Procin ® Laboratorios Pisa) Ketamina (Anesket® Laboratorios Pisa) en una dilución 0.8:0.2, aplicado de forma intravenosa en la vena yugular. Posteriormente la oveja se preparó en la camilla colocada en un plano a 45°, se aplicó 1 ml de lidocaína (Pisacaina ® Laboratorios Pisa). Se colocaron los campos quirúrgicos y se procedió a hacer una incisión de 7 cm sobre la línea media ventral y con unas pinzas de Bab-cock exponer el aparato reproductor. Con ayuda de un catéter intravenoso (Punzocat ® de 32 mm) se hizo una punción en la unión útero-tubárica para ingresar al lumen del cuerno uterino; enseguida se realizó una segunda punción a nivel del ligamento intercornual para introducir un catéter Foley No. 10. Una jeringa con 60 ml de un medio de lavado (Vigro Complete Flush Solution, Laboratorios Bioniche®) a 37 °C fue conectada al catéter intravenoso para que por arrastre los embriones fueran recolectados en cajas Petri cuadriculadas con la solución que salía por el extremo anterior de la sonda Foley. Las cajas de Petri se trasladaron al laboratorio para iniciar la búsqueda y evaluación de los embriones.
La búsqueda de los embriones se realizó con un microscopio estereoscópico (Barnstead, USA) a un aumento de 40X, de manera minuciosa, observando cada uno de los cuadrantes de la caja de Petri. Las estructuras encontradas se colocaron en una caja Petri de cuatro pozos que contenía 200 µl de medio de mantenimiento (Holding®, Laboratorios Bioniche) en cada pozo, colocadas en una platina térmica a 37 °C. Los embriones fueron evaluados y clasificados en el microscopio estereoscópico a un aumento de 100X, de acuerdo a su morfología, y en base a los criterios de la Sociedad Internacional de Transferencia de Embriones (IETS por sus siglas en inglés(13)).
Los embriones se transfirieron en fresco, seis días y medio posteriores a la aparición del estro (tratando de aminorar el tiempo de asincronía entre la donadora y la receptora), seleccionando hembras tanto del grupo DB+AM como del grupo DB-AM, asegurándose de que a todas las hembras de los cuatro tratamientos, les tocará el mismo número y calidad de embrión transferido. Con un periodo previo de 24 h de ayuno, la receptora se colocó en una camilla y se llevó a cabo el mismo protocolo que para la colecta de embriones. Se visualizaron los cuernos uterinos por laparoscopia y mediante una laparotomía media ventral, se realizó una incisión de 2 cm en la línea media abdominal, con una pinza Bab-cock se exteriorizó el cuerno uterino ipsilateral del ovario con uno o más cuerpos lúteos normales, hidratándolo permanentemente con solución salina fisiológica (Solución CS® Pisa Laboratorios). Con ayuda de un catéter intravenoso (Punzocat ® de 32 mm) se hizo una punción en la unión útero-tubárica, se colocó un embrión de calidad 1 o 2 (dependiendo la disponibilidad) en un catéter (TOMCAT®) bajo el siguiente orden: una columna de medio, aire, medio con dos embriones, aire y finalmente medio. Con una jeringa de insulina, los embriones colocados en el catéter se impulsaron hacia la luz del cuerno uterino. La sutura de los planos quirúrgicos se realizó con Catgut® crómico calibre cero para peritoneo y el músculo, y la piel se suturó con Nylon calibre cero. Para prevenir infecciones post-operatorias y una vez terminada la sutura se aplicaron 100 mg de oxitetraciclina IM (Emicina LA ®, Laboratorios Pfizer).
Las variables evaluadas fueron:
Manifestación de estro (%): porcentaje de ovejas que mostraron signos de estro dentro de cada tratamiento.
Tiempo a la manifestación del estro (h): horas transcurridas desde el retiro del dispositivo hasta la presentación del estro.
Tasa ovulatoria (TOV): número de cuerpos lúteos presentes al momento de la extracción de los embriones (6.5 días después del estro).
Embriones calidad 1 (EQ1): número de mórulas o blastocistos calidad 1.
Embriones calidad 2 (EQ2): número de mórulas o blastocistos calidad 2.
Embriones transferibles (ET): número de mórulas o blastocistos calidad 1 y 2.
Ovocitos (OV): número de ovocitos que no fueron fertilizados.
Estructuras totales (ST): número de estructuras colectadas totales.
Porcentaje de gestación: número de ovejas receptoras gestantes a los 40 días post transferencia entre el número de ovejas en cada tratamiento, por 100.
Tasa de parición: número de ovejas receptoras paridas en relación al número de ovejas transferidas en cada tratamiento.
La respuesta en las ovejas donadoras se analizó utilizando una regresión de Poisson mediante el Modelo Lineal Generalizado (PROC GENMOD). Para analizar el porcentaje de gestación en las receptoras, se utilizó el análisis de tablas de contingencia para dos factores (DAM y RAM) utilizando el modelo Log lineal a través del procedimiento CADMOD del paquete estadístico SAS(14). La manifestación del estro se analizó mediante una prueba de independencia de Ji-cuadrada utilizando el procedimiento PROC FREC y el tiempo a la manifestación del estro fue analizada con el método de curvas de supervivencia de Kaplan Meier usando el procedimiento LIFETEST y se establecieron diferencias entre tratamientos con la prueba de LOG RANK.
Resultados
La proporción de ovejas donadoras con signos de estro y el tiempo al estro fueron similares entre tratamientos (P>0.05); sin embargo, la inclusión de AM en la dieta de ovejas donadoras incrementó (P<0.05) la TOV, ET, EQ1, OV, y la cantidad de ST por oveja (Cuadro 2).
Cuadro 2: Respuesta de ovejas donadoras Pelibuey alimentadas con dieta base con aceite de maíz (DB+AM) o con dieta base sin aceite de maíz (DB-AM) (Medias ± EE)
ab Valores con diferente literal dentro de hilera presentan diferencias (P<0.05).
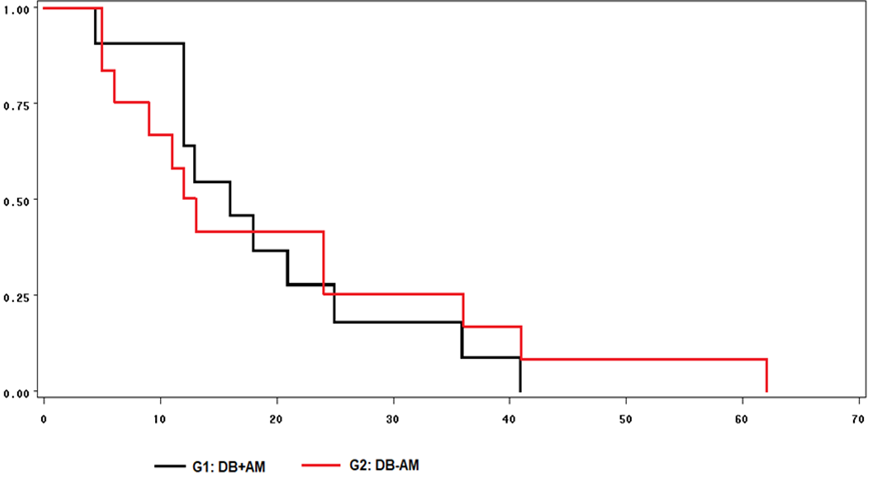
No se observaron diferencias en la probabilidad de manifestación de estro entre grupos, obtenidas por medio de las curvas de supervivencia (Figura 1). Así, el 100 % de las ovejas con DB+AM manifestaron estro a las 42 h post retiro del CIDR mientras que las ovejas DB-AM lo hicieron hasta las 62 h.
Figura 1: Curvas de supervivencia para el tiempo transcurrido entre el retiro del CIDR y la manifestación del estro en ovejas donadoras Pelibuey alimentadas con dieta base con aceite de maíz (DB+AM) o con dieta base sin aceite de maíz (DB-AM)
Respuesta en receptoras
La inclusión de AM en la dieta no alteró el tiempo ni el número de ovejas receptoras en estro
con dieta base con aceite de maíz (DB+AM, 24.08 ± 1.03) o con dieta base sin aceite de maíz (DB-AM, 25.8 ± 3.0).
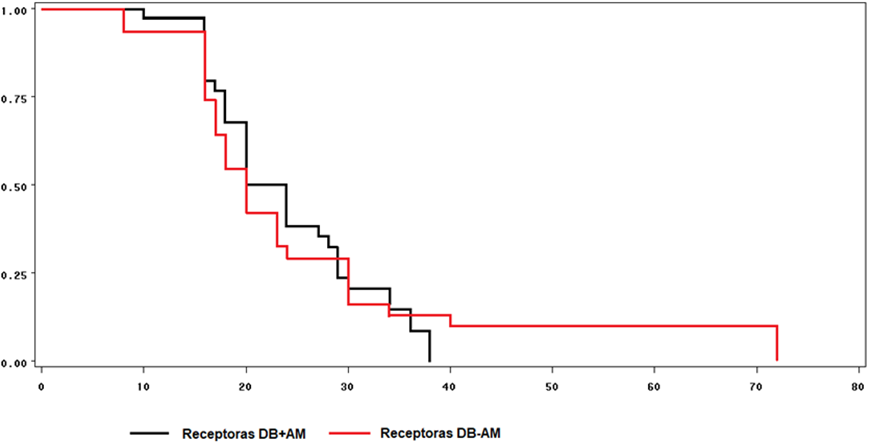
Las curvas de supervivencia para la manifestación del estro en receptoras no mostraron diferencias entre tratamientos (Figura 2). El 100% de las ovejas con AM manifestaron estro a las 38 h post retiro del CIDR mientras que las ovejas testigo lo hicieron a las 72 h.
Figura 2: Curvas de supervivencia para el tiempo transcurrido entre el retiro del CIDR y la manifestación del estro en ovejas receptoras Pelibuey alimentadas con dieta base con aceite de maíz (DB+AM) o con dieta base sin aceite de maíz (DB-AM)
No se encontraron diferencias (P>0.05) ni en el porcentaje de gestación y parición para ninguno de los tratamientos de las receptoras (Cuadro 3).
Cuadro 3: Respuesta en ovejas receptoras Pelibuey alimentadas con dieta base con aceite de maíz (DB+AM) o con dieta base sin aceite de maíz (DB-AM)
(P>0.05).
Discusión
La respuesta a la manifestación y el tiempo al estro en ovejas Pelibuey donadoras y receptoras, no presentó diferencias entre tratamientos (P>0.05). En forma similar, en una investigación con ovejas Corriedale, se reportó un 100 % de ovejas superovuladas en estro con eCG y FSH durante la época de cría(15). En el presente estudio el 25 % de las ovejas donadoras no respondió al protocolo de superovulación, lo cual coincide con otros trabajos en los que la proporción de hembras que no respondieron a la superovulación fue entre 20(16) y 30 % (17). Al suplementar AGP omega 6 (n-6) se incrementa el consumo de energía, ya que, estos son esenciales para la alimentación en rumiantes, además de que participan en los procesos de foliculogénesis, ovulación y conducta estral(18). La población de folículos medianos y grandes se incrementó cuando se suplementó aceite de maíz como fuente de AGP en ovejas, lo que puede dar lugar a una mayor producción de estrógenos por parte de esos folículos, ejerciendo un efecto similar a la eCG(19). En el presente estudio la inclusión de AGP en la dieta de las ovejas no modificó su comportamiento reproductivo, posiblemente debido al corto tiempo de exposición a los AGP en la dieta; ya que se ha mencionado, que para observar una respuesta en la conducta reproductiva de las hembras, se requiere de una exposición de al menos 20 días antes del día del empadre y continuar durante el empadre(20).
El tiempo a la manifestación de estro entre las receptoras (24.08 vs 25.8 h) y donadoras (19.13 vs 20.6 h) fue similar entre tratamientos (P>0.05). Resultados de otros autores mostraron un inicio de estro de 34.5 ± 2.6 h(21) o de 32 ± 5.6 h(22) cuando se aplicaron 500 UI de eCG, difiriendo de lo encontrado en esta investigación. Se ha mencionado que el tiempo al inicio del estro está determinado por la raza y el tipo de FSH empleados en los protocolos de supereovulación(23). El tiempo al inicio del estro fue 24 h más corto cuando las ovejas fueron sometidas a una infusión intravenosa de aceite de oliva comparado con aceite de soya. Se ha mencionado que el aceite de oliva, estimula las concentraciones plasmáticas de metabolito alfa F2 de prostaglandina (PGFM) y prostaglandina E2 (PGE2)(24) aportando evidencia de la posible relación entre los ácidos grasos, las prostaglandinas y el inicio del estro(25). En la presente investigación no fue posible observar tal efecto, probablemente debido a las hormonas utilizadas para la sincronización.
La inclusión de AM en la dieta de ovejas donadoras mejoró la respuesta a la superestimulación en el G1, resultados similares a los encontrados por Herrera-Camacho(6) al evaluar ovejas Pelibuey superovuladas y suplementadas con AM (4% MS), observándose un incremento en el número de cuerpos lúteos, embriones colectados y mórulas; por su parte, Zeron(26) reportaron una mayor calidad en los ovocitos y un incremento en la proporción de AGP en plasma, líquido folicular y células del cumulus ooforus, al suplementar ovejas con jabones cálcicos de aceite de pescado. La nutrición durante la maduración de los ovocitos tiene efectos importantes en la viabilidad de los embriones en ovejas superovuladas(27). Los AG pueden afectar directamente la maduración del ovocito a través de la composición de su membrana(28), o indirectamente afectando la concentración de metabolitos en el líquido folicular, impactando en su posterior desarrollo y viabilidad(29). Por tanto, se sugiere que la inclusión de AM en esta investigación pudo influir en la calidad de los ovocitos y de los embriones durante las primeras etapas de desarrollo, ya que se obtuvieron más embriones transferibles cuando se adicionó AGP en la dieta; además, el incremento en el número de cuerpos lúteos pudo haber modificado la secreción de progesterona, al tener una mayor disponibilidad de colesterol para el tejido lúteo(6), optimizando las condiciones para un mejor desarrollo embrionario.
No se observaron diferencias en el porcentaje de gestación entre los cuatro tratamientos (P>0.05). Resultados similares fueron encontrados en otras investigaciones en donde al adicionar AM en la dieta de ovejas Pelibuey no observaron diferencias en el porcentaje de gestación(7,19). Al respecto se ha mencionado que tanto la subnutrición como la sobrealimentación pueden provocar alteraciones en el medio ambiente intrauterino que impiden un adecuado desarrollo del embrión, afectando su viabilidad(30).
Las dietas que contienen altas concentraciones de AGP como el ácido linoleico en el aceite de maíz, pueden regular la síntesis de prostaglandinas y sus efectos posteriores sobre la persistencia del cuerpo lúteo, síntesis de E2(4,8,25) y el reconocimiento materno de la preñez(31). Además los AGP pueden incrementar las concentraciones circulantes de progesterona, debido a un aumento en la disponibilidad de colesterol(32), el principal precursor para la síntesis de progesterona (P4) en el cuerpo lúteo(33).
La respuesta a la superovulación encontrada en el presente estudio fue similar a lo reportado en otras investigaciones, obteniendo un incremento en la cantidad de embriones transferibles en las donadoras que recibieron los AGP, sin embargo, la adición de AGP en la dieta de ovejas receptoras no mejoró el porcentaje de gestación, por lo cual es necesario realizar más estudios sobre el tiempo de exposición de las hembras a la dieta con AGP, así como la cantidad y calidad de los AGP que se ofrecen, ya que en ovinos, existen muy pocos estudios publicados donde se expliquen los efectos directos de los ácidos grasos poliinsaturados sobre la fertilidad(25).
Conclusiones e implicaciones
En conclusión, las ovejas donadoras presentan buena respuesta a la inclusión de aceite de maíz en un programa de transferencia de embriones. El uso de aceite de maíz en la dieta de ovejas Pelibuey incrementa la tasa ovulatoria, el número de estructuras totales y embriones transferibles, pero no tiene efecto en el porcentaje de preñez de las hembras receptoras de embriones.
Agradecimientos
Al Laboratorio de Reproducción de Ovinos y Caprinos (LaROCa), Campus Montecillo del Colegio de Postgraduados y a la Líneas de Generación y Aplicación del Conocimiento (LGAC) Innovación Tecnológica y Seguridad Alimentaria en Ganadería por su apoyo financiero.
Literatura citada: