https://doi.org/10.22319/rmcp.v15i4.6440
Article
In vitro anthelmintic evaluation of curcumin against the eggs and larvae of three Haemonchus contortus isolates with different susceptibility to ivermectin
Lisandro Alberto Encalada-Mena a
Raquel López-Arellano b
María Eugenia López-Arellano c
María Gabriela Mancilla-Montelongo d
Carlos Alfredo Sandoval-Castro e*
Agustín Olmedo-Juárez c
Juan Felipe de Jesús Torres-Acosta e
a Universidad Autónoma de Campeche, Facultad de Ciencias Agropecuarias, Campeche, México.
b Universidad Nacional Autónoma de México. FES-Cuautitlán, Laboratorio de Ensayos de Desarrollo Farmacéutico, Estado de México, México.
c Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Centro Nacional de Investigación Disciplinaria en Salud Animal e Inocuidad, Morelos, México.
d CONAHCYT-Universidad Autónoma de Yucatán. Facultad de Medicina Veterinaria y Zootecnia, Yucatán, México.
e Universidad Autónoma de Yucatán, Facultad de Medicina Veterinaria y Zootecnia, Km 15.5 Carr. Mérida-Xmatkuil, 97315, Mérida, Yucatán, México.
*Corresponding author: carlos.sandoval@correo.uady.mx
Abstract:
Increasing gastrointestinal nematode resistance to conventional anthelmintics (AH) is a growing worldwide problem. Among various sources, the search for alternative AH has focused on plant secondary metabolites such as curcumin. The in vitro AH activity of curcumin (CUR) was evaluated against three isolates of the ruminant nematode Haemonchus contortus with different susceptibilities to conventional AH. Four in vitro tests were run: egg hatching inhibition (EHI), larval migration inhibition (LMI), larval exsheatment inhibition (LEI) and 72 h mortality of unsheathed L3. Curcumin (CUR) concentration range was 0 - 8.5 μg CUR/mL in the EHI, LMI and mortality tests. In the LEI test it was 0 - 17.3 μg CUR/mL. Concentration-response curves were generated using a log-logistic regression. Experimental design was completely random and results were analyzed with an ANOVA. Curcumin did not exhibit AH activity in the EHI, LMI and mortality tests, but had a significant AH effect in the LEI test. This effect was strongest against the FMVZ-UADY isolate (EC50= 1.9 μg/mL, 95%CI= 1.58-2.31), followed by the Paraíso isolate (EC50= 3.2 μg/mL, 95%CI = 2.69-3.81) and the CENID-SAI, INIFAP isolate (EC50= 7.0 μg/mL; 95%CI= 6.58-7.43). At the evaluated doses, curcumin had an AH effect against exsheatment of H. contortus L3, but no effect on egg hatching, L3 migration or mortality of exsheathed L3.
Key words: Polyphenol, Anthelmintic, Larval exsheathed, Polymeric stabilizers.
Received: 26/04/2023
Accepted: 03/07/2024
Introduction
Gastrointestinal nematodes (GIN) are a major health problem in grazing ruminants(1,2). Small ruminants with high GIN infections suffer digestive disorders that cause growth retardation or decrease productivity parameters. These parasites can also cause anemia, edema and, in very severe cases, death(3,4). Haemonchus contortus is one of the most important GIN due to its pathogenicity and worldwide distribution in tropical and subtropical regions(5).
Control of H. contortus is based on commercial dewormers that can, over time, select for populations with anthelmintic resistance (AR)(6). Increasing AR is the driving force behind the search for alternative helminth control measures(7). Plant secondary metabolites, particularly polyphenolic compounds, have an anthelmintic (AH) effect against different H. contortus life stages(8,9). Turmeric Curcuma longa L., a member of the Zingiberaceae family and native to Asia, contains polyphenols. The main polyphenol in C. longa extracts is curcumin (CUR) (60-75 %), although smaller proportions of desmethoxycurcumin and bisdesmethoxycurcumin are also present(10,11). Curcumin has confirmed pharmacological activities such as antioxidant, anti-inflammatory, anticancer, antiviral, antibacterial, and antiparasitic(12-18). Its antiparasitic activity has been evaluated using extracts produced with solvents of different polarity, and different elements of C. longa plants. These extracts’ AH activity has been evaluated using different concentrations (mg/mL) against H. contortus adults, L3 and eggs(19-21). High doses are used because CUR exhibits low hydrosolubility, poor absorption, and rapid degradation, which reduce its bioavailability(10).
Research has focused on increasing CUR solubility by encapsulating secondary metabolites in lipid nanoparticles, nanoemulsions, nanoliposomes, biodegradable polymers and dendrimers, and hydrogels, with the use of casein and cyclodextrins(22,23). A combination of CUR and polymeric stabilizers such as polyvinylpyrrolidone (PVP) in a solid dispersion (CUR/PVP) has recently been proposed(24,25). This combination improves CUR solubility and provides low toxicity in cells and tissues(26-28). Few studies have evaluated the in vitro AH activity of C. longa, and none mention the CUR metabolite concentrations used in the bioassays; indeed, they assume that any observed AH effect is caused by CUR and other curcuminoids(19-21). Therefore, the AH activity of CUR against H. contortus has yet to be unequivocally demonstrated. The present study objective was to evaluate the in vitro AH activity of CUR against three H. contortus isolates with different AR status.
Material and methods
Experimental ethics
All experimental animals were handled following applicable laws for germplasm collection (NOM-051-ZOO-1995 and NOM-062-ZOO-1999) (www.gob.mx/senasica). The experimental design and procedures followed the ethical guidelines of the Bioethics Committee of the Faculty of Veterinary Medicine and Zootechny of the Autonomous University of Yucatan (Facultad de Medicina Veterinaria y Zootecnia de la Universidad Autónoma de Yucatán - FMVZ-UADY) (Permit No. CB-CCBA-D-2021-005).
Study area
Production of H. contortus in donor animals with monospecific infections, and all in vitro bioassays, were performed at the FMVZ-UADY, Xmatkuil, Mérida, Mexico.
Haemonchus contortus isolates
Three H. contortus isolates were used in the tests. The AH resistance status of each was determined before their use.
(1) The Paraíso isolate originated from a commercial sheep farm in Umán, Yucatan, Mexico. It is reported to exhibit resistance to ivermectin (IVM; fecal egg count reduction: 64 %), albendazole sulfoxide (ABZ; fecal egg count reduction: 0%), and levamisole (LEV; fecal egg count reduction: 92 %)(29);
(2) The FMVZ-UADY isolate originated from a farm in Merida, Yucatan, Mexico. It has been reported as resistant to ABZ (fecal egg count reduction: 89 %) and LEV (fecal egg count reduction: 87 %)(30), but susceptible to IVM (fecal egg count reduction: 99 %)(29);
(3) The CENID-SAI-INIFAP isolate was provided by the Helminth Department of the National Center for Disciplinary Research in Animal Health and Food Safety (Centro Nacional de Investigación Disciplinaria en Salud Animal e Inocuidad - CENID-SAI) of the National Institute of Forestry, Agriculture and Livestock Research (Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias - INIFAP). In vitro resistance assays were performed at the FMVZ-UADY. Based on the egg hatch methodology described by Von Samson-Himmelstjerna et al(31), this isolate was found susceptible to thiabendazole (TBZ) (EC50= 0.050 μg/mL), and using the larval migration technique(32) it was found susceptible to IVM (EC50= 1.09 μM)(33). In a separate study using the in vitro mortality technique, it was found susceptible to IVM (79.22 % mortality at 11.42 mM)(34).
Donors and collection of monospecific Haemonchus contortus isolates
Donor animals were six goats aged 3 to 4 months, 15 kg live weight (LW), raised GIN-free from birth. For each H. contortus isolate, two donor animals were infected with 7,000 L3 per os(35). At all times, the animals were kept in individual cages with raised floors located inside individual pens with concrete floors at the FMVZ-UADY facilities.
Beginning at 24 d post-infection, fecal samples were collected directly from the rectum of each donor animal in new polyethylene bags(36). The fecal samples were processed following the McMaster technique to quantify the number of eggs H. contortus eggs and quantify them as eggs per gram of feces (EPG)(37). Each test was performed with 2 g of feces and 28 mL saturated sugar solution (density = 1.28, 50 EPG sensitivity). Third stage larvae (L3) were obtained from stool cultures maintained in clean plastic jars incubated at 28 °C for 5 to 6 days. The larvae were recovered using the Baermann technique(38), and identified as Haemonchus using morphological keys(39).
Haemonchus contortus egg collection
Egg collection was done following the MAFF (1986) procedure(38). Feces were collected directly from the rectum of donor animals using polyethylene bags. In the laboratory, the feces were placed in a plastic strainer over a porcelain mortar. For every 10 g of feces, 100 mL water was added and the mixture macerated. The feces/water suspension was filtered through a double layer of gauze placed in a funnel, and the liquid recovered in a flask. This liquid was filtered through a 25 μm mesh with a minimal amount of dechlorinated water. Eggs were recovered from the mesh and placed in 50 mL tubes (5810R, Eppendorf, Germany). These were centrifuged at 453 g for 5 min, and the supernatant removed. A saturated sugar solution (25 mL, density= 1.28) was added to the sediment, The latter was resuspended in a vortex mixer and the tubes centrifuged again. The surface layer of the solution was recovered with a bacteriological loop and placed in another plastic tube containing water purified by reverse osmosis. This procedure was repeated several times to recover the largest possible number of eggs. Concentration per milliliter was estimated and adjusted until attaining a 200 eggs/mL suspension.
Haemonchus contortus L3 production
Third stage larvae (L3) were produced in feces collected every 24 h in polyethylene bags from a mesh placed under each cage. In the laboratory, the feces were washed with running water to remove detritus, and the recovered washed feces were used to perform separate coprocultures for each isolate with a Baermann apparatus(38), and placed in culture bottles with ventilated lids. The bottles were identified with harvest date, isolate name and concentration (L3/mL). The larvae were stored under refrigeration (6-10 °C) until used in the in vitro migration and exsheathing inhibition tests, and the exsheathed L3 larvae mortality test.
Curcumin dispersion in polyvinylpyrrolidone (CUR/PVP)
Curcumin has poor solubility(16,40). Therefore, prior to the in vitro studies, an established procedure was applied to improve turmeric E solubility and absorption(25). Turmeric E (52.28% CUR; Laboratorios Mixim S.A. de C.V., Naucalpan, Mexico) was combined with PVP K30 (Agrimer K-30 Ashland, Columbus, Ohio, U.S.) at a 1:7 ratio. This produced a turmeric E dispersion with a 6.2 % final CUR concentration (CUR/PVP). This procedure was done at the Pharmaceutical Development Testing Laboratory (LEDEFAR), Cuautitlán Higher Education Faculty, National Autonomous University of Mexico (Universidad Nacional Autónoma de México – UNAM).
Preparation of CUR/PVP dispersion stock solutions
For the egg hatching inhibition (EHI), larval migration inhibition (LMI) and 72 h mortality tests of exsheathed L3, a stock solution was prepared with 32.4 mg CUR/PVP dispersion in 20 mL purified water and stirred with a magnetic bar for 2 h. This suspension was centrifuged (BHG, Germany) at 1.057 g for 5 min and the supernatant used in the different bioassay concentrations. For the larval exsheathing inhibition (LEI) test, a stock solution was prepared with 64.8 mg CUR/PVP dispersion in 20 mL purified water.
Curcumin concentration
Because of CUR’s low solubility, real CUR content was quantified in the supernatant suspension used in the bioassays. Suspension CUR concentration was measured following methodologies described in Buchi note No. 747(41) and FSSAI(42). Briefly, the supernatant was analyzed with a UV/VIS spectrophotometer (Lambda 25, Perkin Elmer, Beaconsfield, UK) at 425 nm transmittance. A calibration curve (0 - 6 mg/L) of CUR (Sigma Aldrich® standard cat. C7727, 91% purity) in ethanol was prepared. Pure ethanol (Sigma Aldrich® cat. E7148) was used to correct the background reading. The stock solution prepared with 32.4 mg CUR/PVP was found to contain 11.3 μg CUR/mL, and the stock solution prepared with 64.8 mg CUR/PVP contained 23 μg CUR/mL.
Egg hatching inhibition test
The EHI test was run following Coles et al(43). The egg suspension (200 eggs/mL) was evenly distributed in 24-well plates (1 mL per well). Using the CUR/PVP stock solution containing 11.3 μg/mL CUR, serial dilutions (%) were made to four final concentrations in the wells: 2.3 (20 %), 4.0 (35 %), 5.7 (50 %) and 8.5 (75 %) μg/mL CUR in a 2,000 μL final volume. The positive control was 10 µL Lugol’s solution (0.5 % incubated volume), and the negative control was 1,000 µL purified water. The plate was incubated at 28 ºC in a bacteriological oven. After 48 h, egg hatching was stopped by adding 50 µL Lugol’s solution per well. A total of two replicates were performed with three repetitions per concentration. The content of each well was counted using McMaster chambers and a compound microscope (10x). In the bottom of the chamber, per sample counts were done of morulated eggs (ME), eggs containing unhatched larvae (UL) and L1 larvae. The following formula was used:
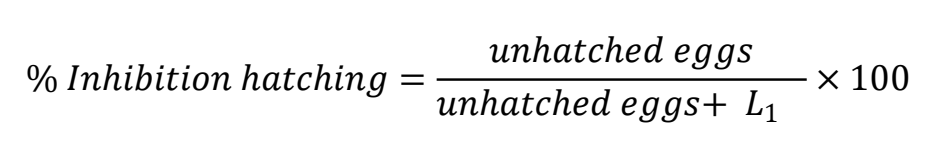
Larval migration inhibition
The LMI test was run to evaluate IVM in dimethyl sulfoxide according to Demeler et al(32), modified as follows for testing with CUR/PVP. The 11.3 μg CUR/mL concentration of CUR/PVP stock solution was used. A serial dilution (%) was done of the stock solution to produce seven final concentrations in the wells: 0, 0.6 (5 %), 1.1 (10 %), 2.0 (17.5 %), 3.7 (32.75 %), 5.7 (50 %) and 8.5 (75 %) μg CUR/mL. In a 24-well plate, 0 - 500 μL purified water plus 0 - 750 μL stock solution were placed in each well. The positive control was 100 μL (10 %) Lugol’s solution, and the negative was 500 μL purified water. In wells with an 8.5 μg CUR/mL final concentration, 250 μL of a larval suspension containing 600 L3/mL was added, with a 1,000 μL final volume. In the remaining wells, 500 μL of a larval suspension containing 300 L3/mL was added, with a 1,000 μL final volume. Two replicates and three repetitions were done for each CUR concentration, and the positive and negative controls. The plates were incubated at 28 °C for 24 h. To prepare the migration plates, 500 μL 1.5 % bacto agar were added to alternating rows, that is, one row with bacto agar and the next empty. A 25 µm mesh was placed in each well containing bacto agar. After incubation in CUR, the L3 content of the working wells and control wells was transferred onto the mesh. The migration plates were incubated at 28 °C for 24 h to allow the L3 to migrate through the mesh. After incubation, the mesh was removed, leaving the migrated L3 larvae in the corresponding wells. Those L3 that did not migrate and remained in the mesh were transferred to the empty wells in the adjacent rows of the same plate. The mesh was washed with 1,000 µL purified water to recover all L3 in the corresponding well. One drop Lugol’s solution was added to all wells, and the contents of each well poured into a McMaster chamber to count L3 per well. The number of migrated and non-migrated L3 in each concentration was counted and percentage of migration calculated using the formula described by Demeler et al(32):
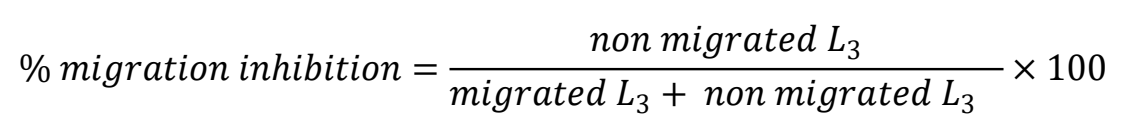
Larval exsheathment inhibition
Inhibition of larval exsheathing (LEI) was quantified following Jackson and Hoste(44). A suspension containing 1,000 L3 per mL, and CUR/PVP dispersion stock solution containing 23.0 μg CUR/mL were used. The L3 were incubated for 3 h at 23 ºC in seven different concentrations, obtained by serial dilution of the stock solution in 15 mL tubes: 0.6 (2.5 %), 2.3 (10 %), 3.5 (15 %), 4.6 (20 %), 8.1 (35 %), 11.5 (50 %) and 17.3 (75 %) μg CUR/mL. Additionally, L3 were incubated in the respective concentrations of PVP K-30 to rule out any AH activity from the polymer. In the positive control, L3 were incubated with LEV (120 mg/mL, Laboratorios Aranda S.A. de C.V., Mexico), and in the negative, they were exposed to purified water. The tubes were centrifuged at 453 g for 5 min and washed with purified water three times. Larvae exposed to the different treatments were divided into four 200 µL aliquots containing 200 L3 each. Prior to the LEI test, calibration curves were generated for each isolate by inducing gradual exsheatment in five dilutions of a sodium hypochlorite (2 %) and sodium chloride (16.5 %) solution (1/300, 1/400, 1/480, 1/600 and 1/800) in phosphate buffer solution (PBS, pH 7.4). Exsheatment was monitored every 20 min (0, 20, 40 and 60 min) in 50 µL aliquots (25-50 L3) using a microscope (10x and 40x). Exsheatment was stopped by flaming the slides covered with coverslips containing the L3. Four replicates were run for each CUR concentration, and %LEI calculated with the formula:
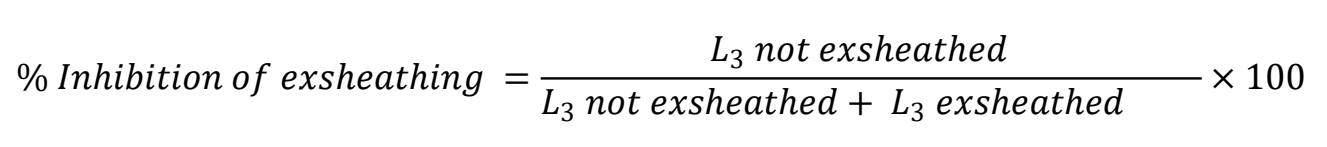
Mortality in exsheathed L3
Mortality in exsheathed L3 caused by CUR was tested following Reyes-Guerrero et al(34), with some modifications. When the L3 of each H. contortus isolate were 3 to 15 wk of age, five mortality trials were run using exsheathed L3 with 0.187 % commercial sodium hypochlorite(45). Using a CUR/PVP dispersion stock solution containing 11.3 μg/mL CUR, exsheathed L3 were exposed to five different concentrations: 1.1 (10 %), 2.3 (20 %), 4.0 (35 %), 5.7 (50 %) and 8.5 (75 %) μg CUR/mL. Lugol’s solution (10 μL) was used as the positive control and purified water (50 μL) as the negative control. Using 96-well microtiter plates, two replicates and three repetitions were done for each CUR concentration, and the positive and negative controls. In wells with an 8.5 μg/mL CUR final concentration, 25 μL containing 100 L3 were added to each well. In the remaining wells, 50 μL containing 100 L3 were added to each well. Final volume in all wells was 100 μL. The plates were incubated at 28 °C for 72 h. After incubation, the live and dead L3 in each well were counted by collecting the entire well in 10 μL drops that were deposited on slides for counting with an optical microscope (4x). Mortality was calculated as a percentage using the formula:
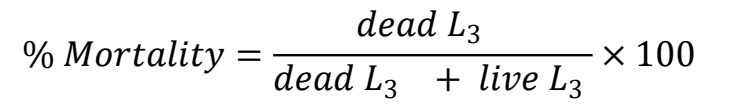
Data analysis
The data produced in the assays involving eggs (EHI) or L3 (LMI, LEI and mortality) were used to generate concentration-response curves in a log-logistic regression with two parameters; the drc extension in the RStudio software was used(46,47). Each test’s EC50 and 95% CI were calculated. The percentage data for EHI, LMI, LEI and mortality were analyzed per test with a completely randomized design, using an ANOVA and the GLM function of the mass extension in the RStudio software(42,43). Two factors were included: the three isolates (Paraíso, FMVZ-UADY and CENID-SAI, INIFAP) and the CUR concentrations in each test. Comparison of the means was run with a Bonferroni test at a P<0.05 significance level.
Before running the ANOVA, a Kolmogorov-Smirnov normality test was run and confirmed that the EHI, LMI and LEI data did not meet the assumption of normality. A Breusch-Pagan heteroscedasticity test found that the same data did not comply with homogeneity of variance. Therefore, the data was transformed with the Box-Cox transformation using the mass extension of the RStudio software(47,48). Analyses were then done using the transformed values, after confirming the assumptions of normality and homogeneity of variance. However, the results are presented transformed to normal values to facilitate interpretation.
Results
CUR inhibition of egg hatching and L1 development
Of the four tested CUR concentrations, only the 5.7 μg/mL CUR achieved 99 % inhibition of hatching in the Paraíso and FMVZ-UADY isolates (Table 1); morulated eggs and larvae were observed (Figure 1 a y b). Inhibition did not decrease significantly for these two isolates even at the highest (8.5 μg CUR/mL) concentration (Table 1). At this same concentration, the L1 exhibited morphological changes that may be associated with larval damage or non-viability (Figure 2 a and b). In the CENID-SAI, INIFAP isolate, the EHI percentages were low (< 10 %) at all concentrations. The EC50 could not be calculated for any of the tested H. contortus isolates.
Table 1: Egg hatching inhibition (%, average ± standard deviation) at five curcumin (CUR) concentrations in three Haemonchus contortus isolates with different susceptibilities to commercial anthelmintics
ab Different letter superscripts in the same column indicate significant difference (P<0.05).
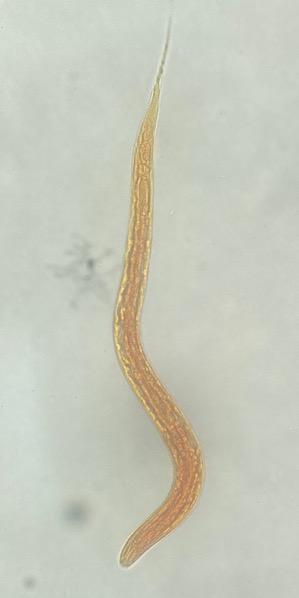
Figure 1: Eggs of the Haemonchus contortus Paraíso isolate (40x) in 5.7 μg/mL CUR concentration in egg hatching inhibition test

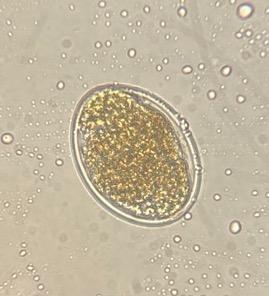
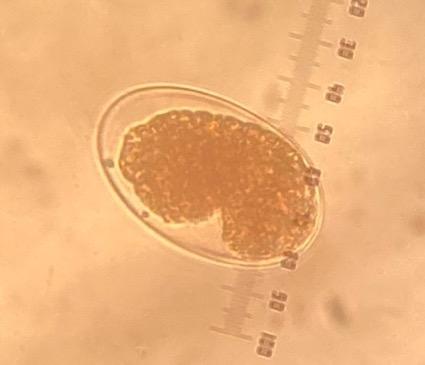
(a) Larvated egg, (b) morulated egg, (c) egg from positive control containing thiabendazole.
Figure 2: L1 larvae of the Haemonchus contortus Paraíso isolate (40x) in 8.5 μg/mL CUR concentration in egg hatching inhibition test
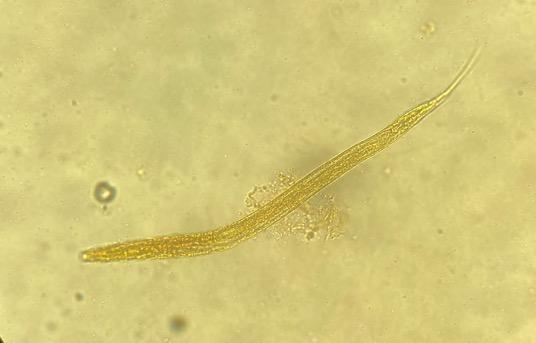
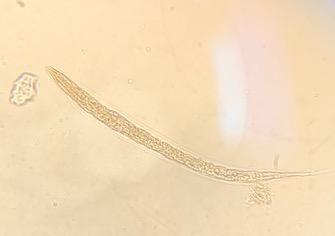
(a) Degraded larva, (b) degraded larva with cuticle separated from body, (c) normal L1 larva in negative control.
CUR inhibition of L3 migration
Of the six tested concentrations, only the 5.7 μg/mL CUR concentration notably inhibited migration compared to the negative control (P<0.05): 64 % inhibition against the Paraíso isolate and 53 % against the FMVZ-UADY isolate (Table 2). The highest inhibition against the CENID SAI, INIFAP isolate was only 20.3 % in the 8.5 μg/mL CUR concentration, which did not differ from the negative control. As occurred with the EHI results, the EC50 could not be calculated for CUR inhibition of migration in the three isolates.
Table 2: L3 migration inhibition (%, average ± standard deviation) at seven curcumin (CUR) concentrations in three Haemonchus contortus isolates with different susceptibilities to commercial anthelmintics
ab Different letter superscripts in the same column indicate significant difference (P<0.05).
CUR inhibition of exsheatment in L3
Exsheatment was inhibited in L3 from 60 to 100 % at 8.1, 11.5 and 17.3 μg CUR/mL. The EC50 of each isolate differed from the others since their 95%CI did not overlap. The lowest values were for the FMVZ-UADY isolate (EC50= 1.9 μg/mL, 95%CI = 1.58-2.31), followed by the Paraíso isolate (EC50= 3.2 μg/mL, 95%CI= 2.69-3.81), and CENID-SAI, INIFAP (EC50= 7.0 μg/mL; 95%CI= 6.58-7.43).
CUR-caused mortality in exsheathed L3
Mortality in exsheathed L3 of all three isolates was less than 10 % at all the tested CUR concentrations. Therefore, CUR had no apparent effect on mortality in exsheathed L3 in these isolates.
Discussion
CUR inhibition of egg hatching
All but one of the tested CUR concentrations had no AH effect on hatching of H. contortus eggs; indeed, no EC50 could be determined for CUR in the three isolates. However, a 99% decrease in hatching was observed with the 5.7 μg CUR/mL concentration in the Paraíso and FMVZ-UADY isolates. Apparently, conditions at this concentration were favorable for solubility of CUR in water and its diffusion through the layers protecting the H. contortus egg. This effect was not observed at the highest concentration (8.5 μg CUR/mL). Why this occurred is unclear. It may be due to high solute saturation in the bioassay liquid, or to a hormetic effect. In the latter, a biphasic response occurs in which low doses have no effect, moderate doses cause change, and high doses exhibit little or none of the expected effect (when graphed it has the shape of an inverted “J” or “U”)(49). This phenomenon is a central theme in the biological, adaptive and repair response, and has implications in pharmacology and toxicology(50). Hormesis is known to occur in different cell types as curcumin concentrations change(51,52). A C. longa methanol:water extract (70:30) is reported to have an AH effect on H. contortus eggs, with an EC50 of 69.75 μg/mL(20). In another study, H. contortus eggs were affected by a hydroalcoholic (1:9) extract of C. longa rhizome (EC50 = 100.9 mg/mL), as well as by an aqueous one (EC50= 83.7 mg/mL)(53). In neither of these two studies was the role of CUR determined in the AH effect of the C. longa extracts against H. contortus eggs. The solvents used in both studies for compound extraction were of varying polarity and the extracts were not purified for identification of the main compounds. Therefore, any effect of curcumin can only be assumed, since this is the main component in C. longa extracts. However, a synergistic effect between different curcuminoids cannot be discounted. For instance, in a study of curcuminoids’ effects on Toxocara canis, CUR alone exhibited an AH effect on L2, but this was augmented when CUR was combined with desmethoxycurcumin, bisdesmethoxycurcumin and cyclocurcumin, suggesting that synergism improved its effectiveness(54).
Of note in the present EHI results is that CUR may cause loss of cellular continuity in L1 at the internal organ level, and these changes may undermine viability in this larval stage. This effect on L1 has not been reported previously, suggesting the need to use the larval development methodology to evaluate the effects of CUR on larvae at different stages(55).
CUR anthelmintic activity against L3
In both the LMI and EHI tests, CUR had no AH effect and the EC50 could not be calculated for any of the three evaluated isolates. As in the EHI test, the best inhibition of L3 migration was with the 5.7 μg CUR/mL concentration, though even at this concentration inhibition percentages were only 50 to 60 %. Results in the EHI and LMI tests were inconsistent, possibly due to degradation of CUR. Indeed, the CUR solution in the wells had changed from a light yellow at the beginning of the bioassay to a brown color at the end (48 h)(56,57). As far as is known, no previous research exists on in vitro LMI testing evaluating any C. longa extract against H. contortus L3.
The most outstanding result is that the evaluated CUR extracts blocked exsheathment in H. contortus L3. This blockage, which is known to occur in ruminal fluid(58), may prevent L3 from invading the crypts in the abomasum, and consequently make larvae incapable of transitioning to later stages, including L4, L5 and adult(59). Blockage of exsheathment is associated with the activity of polyphenols in plant extracts from leaves and other elements(8,60). Curcumin is a polyphenol structurally related to caffeic acid and ferulic acid(61). Both these acids have been evaluated against H. contortus in EHI and LEI bioassays(8). Ferulic acid has ovicidal activity at 200-400 μg/mL, and in the LEI test caffeic acid exhibited activity at 7.8 μg/mL and ferulic acid at 20.6 μg/mL. In the present results, CUR inhibited L3 exsheathment at concentrations < 7 µg/mL, lower than polyphenols mentioned above.
The mechanism by which CUR inhibits exsheathment in the three evaluated H. contortus isolates is unknown. The effectiveness of CUR’s inhibition of exsheathment varied between the three isolates. How effective CUR is at blocking exsheathment may depend on how it interacts with enzymes, proteins, nucleic acids, biomolecules and different receptors types in the sheath of H. contortus(62). A recent metabolomics study reported a panel of metabolites directly responsible for L3 exsheathment, which were associated with amino acids and with the purine and pyruvate metabolism pathways in H. contortus(63). Further research is needed on the possibility that CUR’s effects on one or more of these metabolic pathways is responsible for its inhibition of exsheathment.
The present results confirm that CUR does not affect mortality in exsheathment L3. The low mortality observed here in exsheathment L3 (<10 %) coincides with the low mortality (<10 %) reported elsewhere with a C. longa extract at 100 μg/mL(20), and the absence of any effect on L3 mortality with two C. longa extracts(53). These studies contrast with the work of Nasai et al(21), in which an ethanolic C. longa extract increased L3 mortality (78 %) at doses of 200 mg/mL. However, a large amount of extract was used in this study, which is unfeasible under in vivo conditions. Nonetheless, even though no EC50 could be estimated for CUR in the L3 mortality assay, the EHI test demonstrated that CUR can cause changes compatible with non-viability of H. contortus L1.
Finally, the results confirm that H. contortus isolates from different geographic regions exhibit different in vitro susceptibility to natural plant compounds, both in the EHI and LEI tests(64,65). These previous studies suggest parasites implement an adaptive process to survive in the presence of secondary compounds with AH activity ingested by the ruminant during grazing. In the present study, the isolates had varying responses at the evaluated CUR concentrations in the different in vitro bioassays. This was most evident in the LEI tests in which the three isolates’ responses differed notably from each other. It is unclear why the CENID-SAI, INIFAP isolate exhibited the lowest sensitivity to the CUR extract concentrations compared to the two isolates from Yucatan.
Conclusions and implications
The evaluated curcumin extract had no in vitro anthelmintic activity against egg hatching, L3 migration or mortality in exsheathed L3 in any of the evaluated H. contortus isolates (FMVZ-UADY, Paraíso, CENID-SAI, INIFAP). However, it did inhibit exsheathment in L3, with variable responses in each of the isolates.
Conflict of interests
The authors declare no conflicts of interest.
Acknowledgements
The research reported here was funded by the CENID SAI, INIFAP through project 20454534898, and forms part of the PhD project of Lisandro Encalada Mena at the FMVZ-UADY. Lisandro Encalada Mena received a graduate studies scholarship from the PRODEP-SEP program (UNACAM-122). Production of H. contortus eggs and larvae of the three isolates was done using material from the FMVZ-UADY GIN ceparium, a product of the Cátedras CONAHCYT No. 692 project.
Literature cited:
- Encalada-Mena LA, Corbala-Bermejo JA, Vargas-Magaña JJ, García-Ramírez MJ, Uicab-Brito L, del Río-Rodríguez J. Gastrointestinal nematode prevalence in dual purpose systems in the municipality of Escárcega, Campeche, México. Agrociencia 2009;43(6):569–576.
- Rinaldi L, Catelan D, Musella V, Cecconi L, Hertzberg H, Torgerson PR. et al. Haemonchus contortus: spatial risk distribution for infection in sheep in Europe. Geospat Health 2015;19:325–331.
- Coop RL, Holmes PH. Nutrition and parasite interaction. Int J Parasitol 1996;26(8-9):951-962.
- Arsenopoulos KV, Fthenakis GC, Katsarou EI, Papadopoulos E. Haemonchosis: A Challenging parasitic infection of sheep and goats. Animals 2021;11:1–28.
- Besier RB, Kahn LP, Sargison ND, Van Wyk JA. The Pathophysiology, Ecology and Epidemiology of Haemonchus contortus Infection in Small Ruminants. Adv Parasitol 2016;93:95-143.
- Rose H, Rinaldi L, Bosco A, Mavrot F, De Waal T, Skuce P. et al. Widespread anthelmintic resistance in European farmed ruminants: a systematic review. Vet Rec 2015;176(21):546-546.
- Torres-Acosta JFJ, Hoste H, Sandoval-Castro CA, Torres-Fajardo RA, Ventura-Cordero J, González-Pech P. et al. The art of war against gastrointestinal nematodes in sheep and goat herds of the tropics. Rev Académica Cienc Anim 2019;17:39-46.
- Mancilla-Montelongo G, Castañeda-Ramírez GS, Torres-Acosta JFJ, Sandoval-Castro CA, Borges-Argáez R. Evaluation of cinnamic acid and six analogues against eggs and larvae of Haemonchus contortus. Vet Parasitol 2019;270:25-30.
- Lima CS, Pereira MH, Gainza YA, Hoste H, Regasini LO, de Souza Chagas AC. Anthelmintic effect of Pterogyne nitens (Fabaceae) on eggs and larvae of Haemonchus contortus: Analyses of structure-activity relationships based on phenolic compounds. Ind Crops Prod 2021;164:113348.
- Nelson KM, Dahlin JL, Bisson J, Graham J, Pauli GF, Walters MA. The essential medicinal chemistry of curcumin. J Med Chem 2017;60(5):1620–1637.
- Tsuda T. Curcumin as a functional food-derived factor: degradation products, metabolites, bioactivity, and future perspectives. Food Funct 2018;9(2):705-714.
- Mimche PN, Taramelli D, Vivas L. The plant-based immunomodulator curcumin as a potential candidate for the development of an adjunctive therapy for cerebral malaria. Malar J 2011;10(Suppl 1):1-9.
- Rauf A, Imran M, Orhan IE, Bawazeer S. Health perspectives of a bioactive compound curcumin: A review. Trends Food Sci Technol 2018;74:33–45.
- Trujillo J, Chirino YI, Molina-Jijón E, Andérica-Romero AC, Tapia E, Pedraza-Chaverrí J. Renoprotective effect of the antioxidant curcumin: Recent findings. Redox Biol 2013;1(1):448–456.
- Xue X, Yu JL, Sun DQ, Zou W, Kong F, Wu J. et al. Curcumin as a multidrug resistance modulator - A quick review. Biomed Prev Nutr 2013;3(2):173–176.
- Mahmood K, Zia KM, Zuber M, Salman M, Anjum MN. Recent developments in curcumin and curcumin based polymeric materials for biomedical applications: A review. Int J Biol Macromol 2015;81:877–890.
- De Aguiar DP, Moscardini MBM, Morais ER, De Paula RG, Ferreira PM, Afonso A. et al. Curcumin generates oxidative stress and induces apoptosis in adult Schistosoma mansoni worms. PLoS One 2016;11(11):1–25.
- Cervantes-Valencia ME, Alcalá-Canto Y, Sumano-Lopez H, Ducoing-Watty AM, Gutierrez-Olvera L. Effects of Curcuma longa dietary inclusion against Eimeria spp. in naturally-infected lambs. Small Ruminant Res 2016;136:27-35.
- Singh A, Varshneya C, Telang RS. In vitro anthelmintic effect of Curcuma longa. Indian Vet J 2005;82(6):594–596.
- Ullah S, Khan MN, Sajid MS, Muhammad G. Comparative anthelmintic efficacy of Curcuma longa, Citrullus colocynthis and Peganum Harmala. Glob Vet 2013;11(5):560–567.
- Nasai NB, Abba Y, Abdullah FFJ, Marimuthu M, Tijjani A, Sadiq MA. et al. In vitro larvicidal effects of ethanolic extract of Curcuma longa Linn on Haemonchus larval stage. Vet World 2016;9(4):417–420.
- Yallapu MM, Jaggi M, Chauhan SC. Curcumin nanoformulations: a future nanomedicine for cancer. Drug Discov 2012;17(1-2):71-80.
- Rafiee Z, Nejatian M, Daeihamed M, Jafari SM. Application of curcumin-loaded nanocarriers for food, drug and cosmetic purposes. Trends Food Sci Technol 2019;88:445-458.
- Paradkar A, Ambike AA, Jadhav BK, Mahadik KR. Characterization of curcumin–PVP solid dispersion obtained by spray drying. Int J Pharm 2004;271(1-2):281-286.
- Hernandez-Patlan D, Solis-Cruz B, Pontin KP, Latorre JD, Baxter MF, Hernandez-Velasco X. et al. Evaluation of a solid dispersion of curcumin with polyvinylpyrrolidone and boric acid against Salmonella enteritidis infection and intestinal permeability in broiler chickens: A pilot study. Front Microbiol 2018;9:1–10.
- Teodorescu M, Bercea M. Poly(vinylpyrrolidone) – A versatile polymer for biomedical and beyond medical applications. Poly Plast Technol Eng 2015;54(9):923–943.
- Sadeghi F, Ashofteh M, Homayouni A, Abbaspour M, Nokhodchi A, Garekani HA. Antisolvent precipitation technique: A very promising approach to crystallize curcumin in presence of polyvinyl pyrrolidon for solubility and dissolution enhancement. Colloids Surf B 2016;147:258-264.
- Guo L, Shi M, Song N, Wan Z, Liu H, Liu L. Anchorage of curcumin onto PVP enhances anti-tumor effect of curcumin. Med Chem Res 2019;28(5):646–656.
- Sepúlveda-Vázquez J, Lara-Del Rio MJ, Vargas-Magaña JJ, Quintal-Franco JA, Alcaraz-Romero RA, Ojeda-Chi MM. et al. Frequency of sheep farms with anthelmintic resistant gastrointestinal nematodes in the Mexican Yucatán peninsula. Vet Parasitol: Reg Stud Rep 2021;24:1–7.
- Mancilla-Montelongo MG, González-Pech PG, Miranda-Miranda DY, Castañeda-Ramírez GS, Encalada-Aguilar PA, Can Celis AC. et al. Targeted treatment strategies for the control of gastrointestinal nematodes in a goat flock with anthelmintic resistant worm populations and poor nutrition. Small Rumin Res 2023;227:107062.
- Von Samson-Himmelstjerna G, Coles GC, Jackson F, Bauer C, Borgsteede F, Cirak VY. et al. Standardization of the egg hatch test for the detection of benzimidazole resistance in parasitic nematodes. Parasitol Res 2009;105(3):825–834.
- Demeler J, Küttler U, von Samson-Himmelstjerna G. Adaptation and evaluation of three different in vitro tests for the detection of resistance to anthelmintics in gastro intestinal nematodes of cattle. Vet Parasitol 2010;170:61–70.
- Encalada-Mena LA, López-Arellano, R, López-Arellano ME, Mancilla-Montelongo M G, Sandoval-Castro CA, Olmedo-Juárez A. et al. Curcumin-PVP improves the in vitro efficacy of ivermectin against resistant and susceptible Haemonchus contortus. Exp Parasitol 2024;256:108670.
- Reyes-Guerrero DE, Cedillo-Borda M, Alonso-Morales RA, Alonso-Díaz MA, Olmedo-Juárez A, Mendoza-de-Gives P. et al. Comparative study of transcription profiles of the P-glycoprotein transporters of two Haemonchus contortus isolates: Susceptible and resistant to ivermectin. Mol Biochem Parasitol 2020;238:111281.
- Ramos-Bruno E, Sandoval-Castro CA, Torres-Acosta JFJ, Sarmiento-Franco LA, Torres-Fajardo R, Chan-Pérez, JI. et al. Nitrogen retention in hair sheep lambs with a gradient of Haemonchus contortus infection. Vet Parasitol 2021;296:109488.
- Mendoza de Gives P, Torres-Acosta JF, Figueroa-Castillo JA, Soberanes-Céspedes N, Mancilla-Montelongo MG, Jasso-Villazul C. et al. Diagnóstico y control sustentable de nematodos gastrointestinales en ovinos y caprinos en la era de la resistencia antihelmíntica. Primera ed. Mérida, Yucatán, México: Editorial Universidad Autónoma de Yucatán; 2023.
- Bauer BU, Pomroy WE, Gueydon J, Gannac S, Scott I, Pfister K. Comparison of the FLOTAC technique with the McMaster method and the Baermann technique to determine counts of Dictyocaulus eckerti L1 and strongylid eggs in faeces of red deer (Cervus elaphus). Parasitol Res 2010;107:555-560.
- Ministry of Agriculture, Fisheries and Food (MAFF). Manual of veterinary parasitological laboratory techniques. 3rd ed. London. Her Majesty´s Stationery Office (HMSO); 1986.
- van Wyk JA, Mayhew E. Morphological identification of parasitic nematode infective larvae of small ruminants and cattle: A practical lab guide. Onderstepoort J Vet Res 2013;80(1):1–14.
- Liu W, Zhai Y, Heng X, Che FY, Chen W, Sun D. et al. Oral bioavailability of curcumin: problems and advancements. J Drug Target 2016;24(8):694–702.
- Buchi (2021) Application note No. 747/2021. Determination of curcuminoids in turmeric powder. https://assets.buchi.com/image/upload/v1623829658/pdf/ShortNotes/SN_747_2021_Determination_of_Curcuminoids_in_turmeric_powder.pdf.
- Food Safety and Standards Authority of India (FSSAI). Manual of methods of analysis of foods: spices, herbs and condiments. New Delhi, Government of India, Ministry of Health and Family Welfare; 2012.
- Coles GC, Jackson F, Pomroy WE, Prichard RK, Von Samson-Himmelstjerna G, Silvestre A. et al. The detection of anthelmintic resistance in nematodes of veterinary importance. Vet Parasitol 2006;136(3–4):167–185.
- Jackson F, Hoste H. In vitro methods for primary screening of plant products for direct activity against ruminant nematodes. In: Vercoe PE, et al editors. In vitro screening of plant resources for extra- nutritional attributes in ruminants: Nuclear and related methodologies. Springer;2010:25–45.
- Liébano-Hernández E, López-Arellano ME, Vázquez-Prats V, Mendoza de Gives P. Cryopreservation of infective larvae of Haemonchus contortus. Rev Latinoam Microbiol 1996;38(2):111-114.
- Ritz C, Baty F, Streibig JC, Gerhard D. Dose-response analysis using R. PLoS One 2015;10(12):1–13.
- R Core Team. A language and environment for statistical computing Vienna, Austria. 2020.
- Ripley B, Venables B, Bates DM, Hornik K, Gebhardt A. Firth D, Ripley MB. Package ‘mass’. Cran R 538. 2013;113-120.
- Calabrese EJ. Hormesis: path and progression to significance. Int J Mol Sci 2018;19(10):2871.
- Calabrese EJ, Mattson MP. How does hormesis impact biology, toxicology, and medicine?. NPJ Aging Mech 2017;3(1):13.
- Moghaddam NSA, Oskouie MN, Butler AE, Petit PX, Barreto GE, Sahebkar A. Hormetic effects of curcumin: What is the evidence? J Cell Physiol 2019;234(7):10060–10071.
- Rainey NE, Moustapha A. Petit PX. Curcumin, a multifaceted hormetic agent, mediates an intricate crosstalk between mitochondrial turnover, autophagy, and apoptosis. Oxid Med Cell Longev 2020;3656419.
- Cruz-Aguila J. Pruebas in vitro de extractos vegetales contra Haemonchus contortus [tesis licenciatura]. México, Tabasco: Universidad Autónoma Chapingo; 2018.
- Kiuchi F, Goto Y, Sugimoto N, Tsuda Y, Akao N, Kondo K. Nematocidal activity of turmeric: synergistic action of curcuminoids. Chem Pharm Bull 1993;41(9):1640–1643.
- Olmedo-Juárez A, Rojo-Rubio R, Arece-García J, Salem AZ, Kholif AE, Morales-Almaraz E. In vitro activity of Pithecellobium dulce and Lysiloma acapulcensis on exogenous development stages of sheep gastrointestinal strongyles. Ital J Anim Sci 2014;13(4):3104.
- Li B, Konecke S, Wegiel LA, Taylor LS, Edgar KJ. Both solubility and chemical stability of curcumin are enhanced by solid dispersion in cellulose derivative matrices. Carbohydr Polym 2013;98(1):1108-1116.
- Rathnayaka RMMP, Madhusankha GDMP, Perera ODAN. Curcumin ethanolic extraction and its’ stability in food applications. In 2019 From Innovation to Impact (FITI), 1-6.
- Marin-Tun CG, Mancilla-Montelongo MG, Capetillo-Leal CM, Sandoval-Castro CA, Hoste H, Borges-Argáez R, Torres-Acosta JFJ. Adapting the in vitro rumen incubation method to evaluate the effect of a plant extract on the exsheathment inhibition of Haemonchus contortus infective larvae. Vet Parasitol 2024;327:110135.
- Sommerville RI. The exsheathing mechanism of nematode infective larvae. Exp Parasitol 1957;6(1):18-30.
- Spiegler V, Liebau E, Hensel A. Medicinal plant extracts and plant-derived polyphenols with anthelmintic activity against intestinal nematodes. Nat Prod Rep 2017;34(6):627-643.
- Hardman WE. Diet components can suppress inflammation and reduce cancer risk. Nutr Res Pract 2014;8(3):233-240.
- Ali R, Rooman M, Mussarat S, Norin S, Ali S, Adnan M, Khan SN. A systematic review on comparative analysis, toxicology, and pharmacology of medicinal plants against Haemonchus contortus. Front Pharmacol 2021;12:644027.
- Palevich N, Maclean PH, Subbaraj AK, Cao M. A multimodal metabolomics approach to elucidate the trigger components of rumen fluid for larval exsheathment in a model gastrointestinal parasitic nematode, Haemonchus contortus. bioRxiv 2021. [preprint]
- Chan-Pérez JI, Torres-Acosta JF, Sandoval-Castro CA, Hoste H, Castañeda-Ramírez GS, Vilarem G. et al. In vitro susceptibility of ten Haemonchus contortus isolates from different geographical origins towards acetone:water extracts of two tannin rich plants. Vet Parasitol 2016;217:53–60.
- Chan-Pérez, JI, Torres-Acosta JFJ, Sandoval- Castro CA, Castañeda-Ramírez GS, Vilarem G, Mathieu C. et al. Susceptibility of ten Haemonchus contortus isolates from different geographical origins towards acetone:water extracts of polyphenol-rich plants. Part 2: Infective L3 larvae. Vet Parasitol 2017;240:11-16.