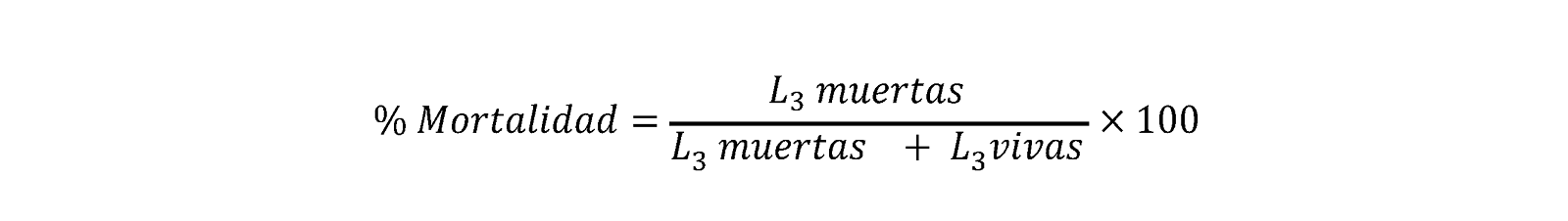https://doi.org/10.22319/rmcp.v15i4.6440
Artículo
Evaluación antihelmíntica in vitro de curcumina contra huevos y larvas de aislados de Haemonchus contortus con distinta susceptibilidad a ivermectina
Lisandro Alberto Encalada-Mena a
Raquel López-Arellano b
María Eugenia López-Arellano c
María Gabriela Mancilla-Montelongo d
Carlos Alfredo Sandoval-Castro e*
Agustín Olmedo-Juárez c
Juan Felipe de Jesús Torres-Acosta e
a Universidad Autónoma de Campeche, Facultad de Ciencias Agropecuarias, Campeche, México.
b Universidad Nacional Autónoma de México. FES-Cuautitlán, Laboratorio de Ensayos de Desarrollo Farmacéutico, Estado de México, México.
c Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Centro Nacional de Investigación Disciplinaria en Salud Animal e Inocuidad, Morelos, México.
d CONAHCYT-Universidad Autónoma de Yucatán. Facultad de Medicina Veterinaria y Zootecnia, Yucatán, México.
e Universidad Autónoma de Yucatán, Facultad de Medicina Veterinaria y Zootecnia, Km 15.5 Carr. Mérida-Xmatkuil, 97315, Mérida, Yucatán, México.
*Autor de correspondencia: carlos.sandoval@correo.uady.mx
Resumen:
El creciente problema de resistencia a los antihelmínticos (AHs) convencionales en poblaciones de nematodos gastrointestinales (NGI), ha motivado la búsqueda de alternativas para su control en los rebaños de ovinos y caprinos, como es el caso de los metabolitos secundarios de plantas. En el presente trabajo se evaluó la actividad AH in vitro de curcumina (CUR) contra tres aislados del nematodo de rumiantes Haemonchus contortus con diferente susceptibilidad a los AHs convencionales. Se utilizaron las pruebas in vitro de inhibición de la eclosión de huevos (IEH), inhibición de la migración larvaria (IML), inhibición del desenvaine larvario (IDL) y mortalidad a las 72 h de L3 sin vaina. Las pruebas se realizaron usando un intervalo de concentraciones de 0 - 8.5 μg CUR/ml. Solamente la prueba de IDL fue realizada con un intervalo de concentración de 0 - 17.3 μg CUR/ml. Se obtuvieron las curvas de concentración-respuesta en una regresión log-logística y se realizó un diseño completamente al azar mediante un ANOVA. La CUR no presentó actividad AH en las pruebas de IEH, IML y mortalidad, pero sí demostró efecto AH significativo en la prueba de IDL. Este efecto fue más potente contra el aislado FMVZ-UADY (CE50= 1.9 μg/ml, IC95% 1.58-2.31), seguido por el aislado Paraíso (CE50= 3.2 μg/ml, IC95% 2.69-3.81) y CENID-SAI, INIFAP (CE50= 7.0 μg/ml; IC95% 6.58-7.43). Se concluye que la CUR tiene efecto AH contra el desenvaine de las L3 de H. contortus pero no afecta la eclosión de huevos, la migración de L3 o la mortalidad de L3 sin vaina a las dosis evaluadas.
Palabras clave: Polifenol, Antihelmíntico, Desenvaine larvario, Estabilizadores poliméricos.
Recibido: 26/04/2023
Aceptado: 03/07/2024
Introducción
Los nematodos gastrointestinales (NGI) representan uno de los principales problemas de salud de rumiantes en pastoreo(1,2). Los pequeños rumiantes que padecen infecciones altas por NGI presentan alteraciones digestivas que ocasionan retraso del crecimiento o disminuyen los indicadores productivos. También pueden causar anemia, edema y, en casos muy severos, la muerte del animal(3,4). Haemonchus contortus es uno de los NGI de mayor relevancia debido a su patogenicidad y su amplia distribución en regiones tropicales y subtropicales del mundo(5). El control de estos parásitos se basa en el uso de desparasitantes comerciales que, con el tiempo han favorecido la selección de poblaciones con resistencia a los antihelmínticos (RA)(6). La creciente problemática de poblaciones de NGI con RA ha motivado la búsqueda de medidas alternativas de control(7). Entre éstas, se encuentra el uso de metabolitos secundarios de plantas, en particular los compuestos polifenólicos, que han demostrado su efecto antihelmíntico (AH) contra diferentes fases de vida del H. contortus(8,9). La planta Curcuma longa L. pertenece a la familia Zingiberácea y es nativa del continente asiático. El principal polifenol encontrado en los extractos obtenidos de C. longa es la curcumina (CUR) (60-75 %), junto con la desmetoxicurcumina y bisdesmetoxicurcumina en menor proporción(10,11). A la CUR se le han confirmado diversas actividades farmacológicas como anti-oxidante, anti-inflamatorio, anti-cancerígeno, anti-viral, anti-bacterial y anti-parasitario(12-18). La evaluación de la actividad antiparasitaria de CUR se ha realizado mediante extractos obtenidos con solventes de diferente polaridad a partir de distintas partes de plantas de C. longa. La actividad AH ha sido evaluada contra adultos, L3 y huevos de H. contortus a diversas concentraciones (mg/ml)(19-21) generalmente empleando altas dosis debido a que la CUR presenta baja solubilidad en agua, pobre absorción, rápida degradación, y por tanto, presenta una reducida biodisponibilidad(10). Estas características han llevado a la búsqueda de procesos que permitan aumentar su solubilidad mediante la encapsulación de los metabolitos secundarios en nanopartículas lipídicas, nanoemulsiones, nanoliposomas, polímeros biodegradables, dendrímeros, hidrogeles, uso de caseína y ciclodextrinas(22,23). Así, recientemente se ha propuesto una combinación de la CUR con estabilizadores poliméricos como la polivinilpirrolidona (PVP) en forma de dispersión sólida (CUR/PVP)(24,25). Esta combinación mejora la solubilidad de la CUR y mantiene baja su toxicidad en células y tejidos(26-28). Por otro lado, los pocos trabajos que han evaluado la actividad AH in vitro de los extractos de C. longa no mencionan las concentraciones del metabolito CUR en los bioensayos, y solo se supone que el efecto AH reportado se debe al contenido de CUR y otros curcuminoides(19-21). Por lo tanto, la actividad AH de la CUR contra H. contortus no se ha demostrado plenamente. El objetivo del presente estudio fue evaluar la capacidad AH in vitro de la CUR sobre tres aislados de H. contortus con distintos estatus de RA.
Material y métodos
Consideraciones éticas experimentales
Todos los animales experimentales se manejaron siguiendo las Normas Oficiales Mexicanas para colecta de germoplasma (NOM-051-ZOO-1995 y NOM-062-ZOO-1999) (www.gob.mx/senasica). Así mismo, se siguieron los lineamientos éticos del Comité de Bioética de la Facultad de Medicina Veterinaria y Zootecnia de la Universidad Autónoma de Yucatán (FMVZ-UADY) (Permiso No. CB-CCBA-D-2021-005).
Lugar del estudio
La producción de H. contortus en animales donadores con infecciones monoespecíficas controladas, y los bioensayos in vitro se realizaron en la FMVZ-UADY, Xmatkuil, Mérida, México.
Aislados de Haemonchus contortus
Se utilizaron tres aislados de H. contortus cuya caracterización de resistencia a AH se conocía antes de realizar el presente estudio:
(1) El aislado “Paraíso” proveniente de una granja ovina comercial ubicada en Umán, Yucatán, México, con antecedentes de resistencia a ivermectina (IVM; i.e. reducción de huevos en heces: 64 %), sulfóxido de albendazol (ABZ; reducción de huevos en heces: 0 %) y levamisol (LEV; reducción de huevos en heces: 92 %)(29).
(2) El aislado “FMVZ-UADY”, originario de una granja ubicada en Mérida, Yucatán, con antecedentes de NGI resistentes a ABZ (i.e. reducción de huevos en heces: 89 %) y LEV (reducción de huevos en heces: 87 %)(30), pero susceptible a IVM (i.e. reducción de huevos en heces 99 %)(29).
(3) El aislado CENID-SAI-INIFAP, fue proporcionado por el departamento de Helmintología del Centro Nacional de Investigación Disciplinaria en Salud Animal e Inocuidad, Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias. Este aislado fue caracterizado en la FMVZ-UADY con técnicas in vitro como susceptible a tiabendazol (TBZ) (CE50= 0.050 μg/ml) de acuerdo con la metodología de eclosión de huevos descrita por Von Samson-Himmelstjerna et al(31), y de acuerdo a la técnica de migración larvaria(32) susceptible a IVM (CE50= 1.09 μM)(33). Así mismo en 2020, este aislado fue caracterizado como susceptible a IVM (79.22 % mortalidad a 11.42 mM) mediante la técnica de mortalidad in vitro(34).
Donadores y obtención de aislados monoespecíficos de Haemonchus contortus
Se utilizaron seis caprinos de entre 3 y 4 meses de edad de 15 kg de peso vivo (PV), criados libres de NGI desde el nacimiento. Para cada aislado, anteriormente descritos, de H. contortus se infectaron dos animales donadores, cada uno recibió 7,000 L3 per os(35). En todo momento los animales se mantuvieron en jaulas individuales con piso elevado ubicadas dentro de corrales individuales con piso de concreto en las instalaciones de la FMVZ-UADY.
A partir de los 24 días post-infección se obtuvieron muestras de heces directamente del recto de cada animal donador usando bolsas de polietileno nuevas(36). Las muestras de heces se usaron para determinar la presencia y el número de huevos por gramo de heces (HPG) mediante la técnica de McMaster(37); esta prueba se realiza con 2 g de heces y 28 ml de solución saturada de azúcar (densidad= 1.28 y sensibilidad de 50 HPG). A continuación, se realizaron coprocultivos en frascos de plástico limpios y se incubaron a 28 °C durante 5-6 días para obtener L3, las cuales se recuperaron de las heces mediante la técnica de Baermann(38). Finalmente fueron identificadas como Haemonchus utilizando claves morfológicas(39).
Obtención de huevos de Haemonchus contortus
La obtención de huevos se realizó de acuerdo al procedimiento de MAFF (1986)(38). Se colectaron las heces directamente del recto de los animales donadores usando bolsas de polietileno. En el laboratorio, las heces se depositaron en un colador de plástico colocado sobre un mortero de porcelana. Se añadieron 100 ml de agua por cada 10 g de heces y se procedió a macerar las heces. La suspensión de heces con agua se filtró usando una capa doble de gasa colocada en un embudo, y el líquido se recuperó en un matraz. Este líquido se tamizó a través de una malla de 25 μm con la mínima cantidad de agua sin cloro. Se recuperaron los huevos del tamiz y se colocaron en tubos de 50 ml. Los tubos (5810R, Eppendorf, Alemania) se centrifugaron a 453 g por 5 min. Después se eliminó el sobrenadante conservando el sedimento, al que se añadieron 25 ml de solución saturada de azúcar (densidad= 1.28). El sedimento se volvió a suspender con ayuda de un vórtex y los tubos fueron centrifugados nuevamente. Se recuperó la capa superficial de la solución con un asa bacteriológica, y ésta fue depositada en otro tubo de plástico con una solución de agua purificada por ósmosis inversa. Este procedimiento se realizó varias veces para tratar de recuperar la mayor cantidad de huevos posibles. Posteriormente se estimó su concentración por mililitro y se ajustó hasta lograr una suspensión de 200 huevos/ml.
Producción de L3 de Haemonchus contortus
La producción de L3 consistió en recuperar cada 24 h todas las heces que los animales donadores depositaron en la malla plástica colocada debajo de cada una de las jaulas individuales. Las heces colectadas de cada donador se pasaron a bolsas de polietileno nuevas claramente identificadas para su traslado al laboratorio. Después de la incubación, las L3 se recuperaron con el aparato de Baermann(38). Las larvas se depositaron en botellas de cultivo con tapa ventilada. Los frascos fueron identificados con su fecha de cosecha, nombre de aislado y su concentración de L3/ml. Las larvas se conservaron en refrigeración (6-10 °C) hasta su uso en las pruebas in vitro de inhibición de la migración y desenvaine, así como la de mortalidad de L3 desenvainadas que son descritas posteriormente.
Dispersión de curcumina en polivinilpirrolidona (CUR/PVP)
Previo a los estudios in vitro y considerando la pobre solubilidad de la CUR(16,40), se utilizó un procedimiento descrito previamente para mejorar la solubilidad y absorción de la cúrcuma E(25). El procedimiento consistió en combinar la cúrcuma E (52.28 % CUR; Laboratorios Mixim S.A. de C.V., Naucalpan, México) con PVP K30 (Agrimer K-30 Ashland, Columbus, Ohio, U.S.), en proporción 1:7. Esto permitió obtener una dispersión de cúrcuma E con una concentración final de CUR de 6.2 % (CUR/PVP). Este procedimiento se realizó en el Laboratorio de Ensayos de Desarrollo Farmacéutico (LEDEFAR) de la Facultad de Estudios Superiores Cuautitlán, UNAM.
Preparación de soluciones madre de la dispersión CUR/PVP
Para las pruebas de inhibición de la eclosión de huevos (IEH), inhibición de la migración larvaria (IML) y mortalidad a las 72 h de L3 sin vaina, se elaboró una solución madre con 32.4 mg de dispersión de CUR/PVP en 20 ml de agua purificada y se agitó con barra magnética durante 2 h. Posteriormente, la suspensión se centrifugó (BHG, Alemania) a 1,057 g durante 5 min y se utilizó el sobrenadante para las distintas concentraciones de los bioensayos. Para la prueba de inhibición del desarrollo larvario (IDL) se elaboró una solución madre con 64.8 mg de dispersión de CUR/PVP en 20 ml de agua purificada.
Determinación de la concentración de curcumina
Debido a la baja solubilidad de la CUR, en el sobrenadante utilizado para los bioensayos fue necesario determinar el contenido real en la suspensión. Para determinar la concentración de la suspensión se utilizó las metodologías descritas en la nota de Buchi No. 747(41) y FSSAI(42). Brevemente, el sobrenadante utilizado se analizó con un espectrofotómetro UV/VIS (Lambda 25, Perkin Elmer, Beaconsfield, UK) a 425 nm de transmitancia. Se preparó una curva de calibración (0 - 6 mg/L) de CUR (estándar de Sigma Aldrich® cat. C7727 con 91 % de pureza) en etanol. Se utilizó etanol puro (Sigma Aldrich® cat. E7148) para corregir la lectura de fondo. Este procedimiento permitió determinar que la solución madre preparada con 32.4 mg de CUR/PVP contenía 11.3 μg CUR/ml y la solución madre de 64.8 mg de CUR/PVP contenía 23 μg CUR/ml.
Prueba de inhibición de eclosión de huevos
La prueba IEH se realizó de acuerdo al procedimiento descrito por Coles et al(43). La suspensión de huevos (200 huevos/ml) se distribuyó de manera homogénea en placas de 24 pozos (1 ml por pozo). Para esta prueba se utilizó la solución madre de CUR/PVP que contenía 11.3 μg/ml de CUR. Las concentraciones finales en los pozos se obtuvieron por diluciones seriadas (%) de la solución madre: 2.3 (20 %), 4.0 (35 %), 5.7 (50 %) y 8.5 (75 %) μg/ml de CUR en un volumen final de 2,000 μl. Se usaron 10 µl de solución de Lugol (0.5 % del volumen incubado) como control positivo de la prueba y 1,000 µl de agua purificada como control negativo. La placa antes mencionada se incubó a 28 ºC en una estufa bacteriológica. Después de 48 h se detuvo la eclosión de los huevos mediante la adición de 50 µl de solución de Lugol por pozo. La prueba contó con dos réplicas y tres repeticiones para cada concentración. El contenido de cada pozo se contabilizó con la ayuda de cámaras de McMaster y un microscopio compuesto (objetivo 10x). En el fondo de la cámara se contabilizó el número de huevos morulados (HM), el número de huevos que contenían larvas que no eclosionaron (LNE) y el número de larvas L1 presentes en la muestra, según la siguiente fórmula:
Prueba de inhibición de migración larval
Esta prueba se realizó de acuerdo al procedimiento de Demeler et al(32) para evaluar IVM en dimetilsulfóxido. Para las pruebas con CUR/PVP, se realizaron las modificaciones que se describen a continuación. Se utilizó la solución madre de dispersión de CUR/PVP en agua purificada a una concentración de 11.3 μg CUR/ml. Las concentraciones finales en los pozos fueron a partir de diluciones seriadas (%) de la solución madre: 0, 0.6 (5 %), 1.1 (10 %), 2.0 (17.5 %), 3.7 (32.75 %), 5.7 (50 %) y 8.5 (75 %) μg CUR/ml. En una placa de 24 pozos se colocaron 0 - 500 μl de agua purificada más 0 - 750 μl de la solución madre en cada pozo. Para el control positivo se emplearon 100 μl (10 %) de Lugol, y para el control negativo 500 μl de agua purificada. Posteriormente, se agregaron en los pozos 250 μl (pozos con concentración final de 8.5 μg CUR/ml) o 500 µl (pozos restantes) de una suspensión de larvas a una concentración de 600 y 300 L3/ml, respectivamente, alcanzando un volumen final de 1,000 μl. Cada concentración de CUR, así como controles positivos y negativos contó con con dos réplicas y tres repeticiones. Las placas se incubaron a 28 °C durante 24 h. Para la preparación de las placas de migración se colocaron 500 μl, de bacto-agar al 1.5 % alternando una fila con bacto-agar y otra fila vacía. Se colocó un tamiz (malla de 25 µm) en cada pozo de bacto-agar. Al final de la incubación con CUR, el contenido de L3 en los pozos de trabajo y en los pozos controles, fue transferido a los tamices. Las placas de migración se incubaron a 28 °C durante 24 h para permitir que las L3 migraran a través de los tamices. Finalizado el tiempo de incubación de las placas de migración se retiró el tamiz. De esta manera las L3 migradas permanecieron en los pozos correspondientes. Las L3 que no migraron (quedando dentro de los tamices) fueron transferidas a los pozos vacíos de las filas contiguas de la misma placa. Los tamices se lavaron con 1,000 µl de agua purificada para recuperar todas las L3 en el pozo correspondiente. Se agregó una gota de solución de Lugol en todos los pozos. El contenido de cada pozo se vertió en cámaras McMaster para realizar el conteo de L3 de cada pozo. Se contó el número de L3 migradas y L3 no migradas de cada concentración y se calculó el porcentaje de migración utilizando la fórmula descrita por Demeler et al(32):
Prueba de inhibición del desenvaine larvario
Esta prueba se realizó de acuerdo al procedimiento de Jackson y Hoste(44). Se empleó una suspensión de 1,000 L3 por ml. Para esta prueba se utilizó la solución madre de dispersión de CUR/PVP que contenía 23.0 μg CUR/ml. Las L3 fueron incubadas durante 3 h a 23 ºC en las diferentes concentraciones, obtenidas a partir de diluciones seriadas de la solución madre: 0.6 (2.5 %), 2.3 (10 %), 3.5 (15 %), 4.6 (20 %), 8.1 (35 %), 11.5 (50 %) y 17.3 (75 %) μg CUR/ml en tubos de 15 ml. Adicionalmente, se incubaron L3 en las respectivas concentraciones de PVP K-30 para descartar cualquier actividad AH del polímero. Para el control positivo las L3 se incubaron con LEV (120 mg/ml, Laboratorios Aranda S.A. de C.V., México). En el control negativo, las L3 fueron expuestas a agua purificada. Posteriormente, se centrifugaron a 453 g durante 5 min y lavadas con agua purificada en tres ocasiones. Las larvas expuestas a los diferentes tratamientos se dividieron en cuatro alícuotas de 200 µl (200 L3 en cada alícuota). Previo a la prueba de desenvaine con CUR/PVP, se realizó la respectiva curva de calibración para inducir el desenvaine gradual de cada aislado con diluciones de una solución de hipoclorito de sodio (2 %) y cloruro de sodio (16.5): 1/300, 1/400, 1/480, 1/600 y 1/800 en solución búfer de fosfatos (PBS, pH 7.4). El proceso de desenvaine fue monitoreado cada 20 min (minutos 0, 20, 40 y 60), en respectivas alícuotas de 50 µl (25-50 L3) usando un microscopio con los objetivos de 10x y 40x. El desenvaine fue detenido mediante el flameado de los portaobjetos cubiertos por los cubreobjetos conteniendo las L3. Se efectuaron cuatro repeticiones por cada una de las concentraciones evaluadas de CUR. El %IDL fue determinado de acuerdo con la siguiente fórmula:
Prueba de evaluación de mortalidad sobre L3 desenvainadas
Esta prueba se realizó de acuerdo al procedimiento de Reyes-Guerrero et al(34) con algunas modificaciones. Durante un periodo que abarcó desde las 3 hasta las 15 semanas de edad de las L3 de cada aislado de H. contortus, se realizaron cinco ensayos con L3 desenvainadas con hipoclorito de sodio comercial al 0.187 %(45). Para esta prueba se utilizó la solución madre de dispersión de CUR/PVP que contenía 11.3 μg/ml de CUR. Las L3 desenvainadas fueron expuestas a diferentes concentraciones 1.1 (10 %), 2.3 (20 %), 4.0 (35 %), 5.7 (50 %) y 8.5 (75 %) μg CUR /ml teniendo como control positivo solución de Lugol (10 μl) y control negativo agua purificada (50 μl). Cada concentración de CUR, controles positivos y negativos contó con dos réplicas de tres repeticiones cada una en placas de microtitulación de 96 pozos. Se agregaron 100 L3, contenidas en 25 μl (pozos con concentración final de 8.5 μg/ml CUR) o en 50 μl (resto de los pozos) en un volumen final de 100 μl por pozo. Posteriormente se incubaron a 28 °C durante 72 h. Al final del periodo de incubación, se procedió a realizar el conteo de L3 vivas y muertas en cada pozo, recogiendo la totalidad del pozo en gotas de 10 μl que fueron depositadas en portaobjetos para su conteo utilizando un microscopio óptico (objetivo 4x). Finalmente, se calculó el porcentaje de mortalidad utilizando la siguiente fórmula:
Análisis de datos
Los datos obtenidos en los ensayos con huevos (IEH) o L3 (IML, IDL y mortalidad) se utilizaron para la obtención de las curvas de concentración-respuesta en una regresión log-logística con dos parámetros en el software RStudio, usando la extensión drc(46,47). Se obtuvo la CE50 e IC95% de cada prueba. El porcentaje de IEH, IML, IDL y mortalidad se analizaron respectivamente en un diseño completamente al azar mediante un ANOVA utilizando la función GLM de la extensión mass en el software RStudio(42,43,48). Se incluyeron dos factores: aislado y concentración, el primero con tres niveles (Paraíso, FMVZ-UADY y CENID-SAI, INIFAP) y el segundo las concentraciones utilizadas de CUR para cada prueba. La comparación de medias se realizó con la prueba de Bonferroni en un nivel de significancia P<0.05.
Previo a realizar el ANOVA descrito, se confirmó que los datos de eclosión, migración y desenvaine no cumplían con el supuesto de normalidad de acuerdo al test de normalidad de Kolmogorov-Smirnov. Tampoco cumplían con la homogeneidad de varianza de acuerdo a la prueba de heterocedasticidad de Breusch-Pagan. Por lo tanto, los datos tuvieron que ser transformados mediante la transformación de Box-Cox con la extensión mass del programa mencionado(47,48), posteriormente se realizó el análisis con los valores transformados una vez confirmados los supuestos de normalidad y homogeneidad de varianza. Sin embargo, los resultados se presentan como valores retro-transformados para facilitar su interpretación.
Resultados
Actividad de la CUR para inhibir el desarrollo del huevo y la eclosión de la L1
En el Cuadro 1 se presentan los promedios de inhibición de eclosión de huevos con distintas concentraciones de CUR para tres distintos aislados de H. contortus. De las cuatro distintas concentraciones de CUR utilizadas, solamente la concentración con 5.7 μg/ml de CUR alcanzó 99 % de inhibición de la eclosión en los aislados Paraíso y FMVZ-UADY, lográndose observar presencia de huevos morulados y huevos larvados (Figura 1 a y b). Sin embargo, a la mayor concentración evaluada, la inhibición no disminuyó de manera significativa para esos mismos aislados (Cuadro 1). En esta última concentración (8.5 μg CUR/ml) las L1 mostraron cambios morfológicos que se podrían asociar a daño o inviabilidad de éstas (Figuras 2 a y b). En el aislado CENID-SAI, INIFAP los porcentajes de IEH fueron bajos (< 10 %) en todas las concentraciones utilizadas. Por tanto, no se pudo determinar la CE50 para ninguno de los aislados de H. contortus evaluados.
Cuadro 1: Porcentaje de inhibición (promedio ± desviación estándar) de la eclosión de huevos tratados con distintas concentraciones de curcumina (CUR) en tres aislados de Haemonchus contortus con distinta susceptibilidad a antihelmínticos comerciales
ab Literal diferente en la misma columna indica diferencia significativa entre aislados (P<0.05).
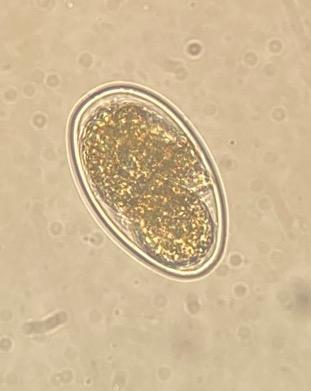
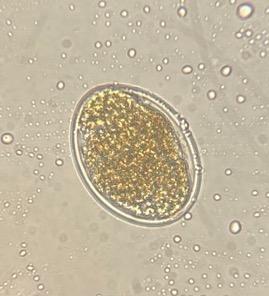
Figura 1: Huevos de Haemonchus contortus aislado Paraíso (objetivo 40x) recuperados del bioensayo de inhibición de la eclosión a una concentración de 5.7 μg/ml de CUR
(a) huevo larvado y (b) huevo morulado. (c) Huevo recuperado del control positivo con tiabendazol.
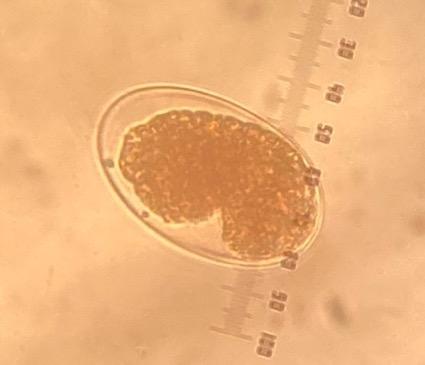
Figura 2: Larvas L1 de Haemonchus contortus aislado Paraíso recuperadas de bioensayos de inhibición de la eclosión de huevos (concentración de 8.5 μg/ml de CUR)
(a) larva degradada y (b) larva degrada con cutícula separada del cuerpo. (c) Larva L1 normal recuperada de control negativo (objetivo 40x).
Actividad antihelmíntica de CUR sobre la migración de L3 de tres aislados de Haemonchus contortus
En el Cuadro 2 se muestran los promedios de IML obtenidos con distintas concentraciones de CUR para tres aislados de H. contortus. De las seis distintas concentraciones utilizadas, solamente la concentración de 5.7 μg/ml de CUR alcanzó porcentajes altos de inhibición de la migración con 64 % y 53 % para los aislados Paraíso y FMVZ-UADY, respectivamente, comparando con el control negativo (P<0.05). Por su parte, el aislado CENID SAI, INIFAP mostró menor inhibición de la migración con solo 20.3 %, y no fue diferente al control negativo. De modo semejante que en los bioensayos de IEH, no fue posible identificar la CE50 de CUR para inhibir la migración de los tres aislados evaluados.
Cuadro 2: Porcentaje de inhibición de migración (promedio ± desviación estándar) de L3 tratadas con distintas concentraciones de curcumina (CUR) en tres aislados de Haemonchus contortus con distinta susceptibilidad a antihelmínticos comerciales
ab Literal diferente en la misma columna indica diferencia significativa entre aislados (P<0.05).
Actividad antihelmíntica de CUR sobre el desenvaine de L3 de tres aislados de Haemonchus contortus
Se demostró que la CUR puede bloquear el desenvaine de las L3 en las concentraciones de 8.1, 11.5 y 17.3 μg CUR/ml en rangos que van desde el 60 % hasta el 100 %. Las CE50 de los tres aislados fueron diferentes al no traslaparse sus respectivos IC95%. La menor CE50 se obtuvo con el aislado FMVZ-UADY (CE50= 1.9 μg/ml, IC95% 1.58-2.31), seguida de Paraíso (CE50= 3.2 μg/ml, IC95% 2.69-3.81) y el valor más alto fue el de CENID-SAI, INIFAP (CE50= 7.0 μg/ml; IC95% 6.58-7.43).
Actividad de CUR sobre la mortalidad de L3 desenvainadas de tres aislados de Haemonchus contortus
Con respecto a los bioensayos de mortalidad de L3 desenvainadas de los tres aislados de H. contortus, los promedios del porcentaje de mortalidad fueron menores al 10 % en todas las concentraciones evaluadas. Por lo tanto, no se registró un efecto de mortalidad sobre las L3 desenvainadas de los tres aislados evaluados.
Discusión
Actividad antihelmíntica de CUR sobre eclosión de huevos de tres aislados de Haemonchus contortus
El presente trabajo demostró que la CUR no tiene efecto AH evidente sobre la eclosión de huevos de H. contortus. No se pudo determinar la CE50 de CUR para ninguno de los aislados evaluados. Sin embargo, es de llamar la atención que se encontró un 99 % de disminución de la eclosión a la concentración de 5.7 μg CUR/ml para los aislados de Paraíso y FMVZ-UADY. Aparentemente, a esta concentración se presentaron las condiciones favorables para la solubilidad de la CUR en el agua y su difusión a través de las capas que protegen al huevo de H. contortus. Sin embargo, en la concentración mayor (8.5 μg CUR/ml) el efecto no fue evidente. Se desconoce por qué se presenta este fenómeno, pero se pudiera deber a la alta saturación del soluto en el líquido del bioensayo, o pudiera ser el resultado de un efecto hormético, al presentarse una respuesta bifásica en la que a bajas dosis no hay efecto y a dosis mayores sí se observan cambios, pero a medida que aumenta la dosis ésta no presenta el efecto esperado (en forma de J o de U invertida al graficarse)(49). Este fenómeno es tema central en la respuesta biológica, adaptativa y de reparación, y tiene implicaciones en farmacología y toxicología(50). Así, la hormesis se ha presentado a distintas concentraciones de curcumina en distintos tipos celulares(51,52). Estudios previos han demostrado que un extracto metanol:agua (70:30) de C. longa mostró efecto AH contra los huevos de H. contortus, con una CE50 de 69.75 μg/ml(20). Por otro lado, Cruz-Aguila(53) demostró el efecto de dos extractos del rizoma de C. longa, uno hidroalcohólico (1:9) y uno acuoso, sobre huevos de H. contortus, con CE50 de 100.9 y 83.7 mg/ml, respectivamente. En los dos estudios que utilizaron extractos de C. longa no se determinó el papel de la CUR en el efecto AH reportado contra los huevos de H. contortus. En esos estudios se utilizaron solventes con distinta polaridad para la extracción de los compuestos y no se purificaron para la identificación de sus principales compuestos. Por lo tanto, estos autores solo suponen que se trata de un efecto de la curcumina, al ser ésta el principal componente en un extracto de C. longa. Sin embargo, es importante considerar la posibilidad de un efecto sinérgico al combinar diferentes curcuminoides como en el caso de Toxocara canis en el que la CUR mostró efecto AH sobre las L2, pero el efecto se incrementó al combinar la CUR con desmetoxicurcumina, bisdesmetoxicurcumina y ciclocurcumina, sugiriendo que el sinergismo mejora la efectividad(54). Es importante recalcar que las L1 observadas en los ensayos de IEH sugieren que la CUR podría ocasionar pérdida de la continuidad celular a nivel de órganos internos, cambios que pueden ser compatibles con la inviabilidad de las L1. Estos efectos en la L1 no habían sido reportados anteriormente y se sugiere que es necesario evaluar el efecto de la CUR sobre larvas de diferente estadio mediante la metodología de desarrollo larvario(55).
Actividad antihelmíntica de CUR sobre L3 de tres aislados de Haemonchus contortus
En las pruebas de IML los resultados son semejantes a los obtenidos en las pruebas de IEH, ya que no se encontró un efecto AH de la CUR y no se pudo determinar la CE50 para ninguno de los tres aislados evaluados. Tal como ocurrió en la prueba de IEH, los mejores porcentajes de inhibición en la migración de L3 se obtuvieron con la concentración de 5.7 μg CUR/ml, aunque solo se alcanzaron porcentajes de inhibición entre 50 y 60 %. Al igual que en las pruebas de IEH, los resultados de la prueba de IML fueron erráticos posiblemente debido a las características propias de la CUR que podría presentar degradación al final de los bioensayos (48 h) ya que fue evidente el obscurecimiento de la solución, pasando de un amarillo claro al inicio del bioensayo hasta un color café en el interior de los pozos a las 48 h(56,57). Hasta donde los autores tienen conocimiento, no existe algún antecedente de pruebas in vitro de IML para evaluar algún extracto de C. longa contra L3 de H. contortus y que pudiera ayudar a explicar los resultados obtenidos.
Un resultado sobresaliente del presente estudio fue encontrar que la CUR bloquea el desenvaine de las L3 de H. contortus. Este bloqueo del desenvaine, que se ha confirmado que ocurre en el líquido ruminal(58), pudiera ocasionar que las L3 no puedan invadir las criptas en el abomaso, y consecuentemente no lograrían llegar a sus siguientes etapas, incluyendo la L4, L5 y adultos(59). Diversos estudios muestran que el bloqueo del desenvaine se asocia a la actividad de los polifenoles contenidos en los extractos de diferentes plantas obtenidos tanto de las hojas como de otras partes de diferentes especies de plantas(8,60). La CUR es un polifenol relacionado estructuralmente con el ácido cafeico y ácido ferúlico(61), los cuales también han sido evaluados contra H. contortus con los bioensayos de IEH e IDL(8). El ácido ferúlico ha mostrado efecto ovicida a concentraciones de 200-400 μg/ml, y en la prueba de IDL los ácidos caféico y ferúlico presentaron actividad a concentraciones de 7.8 y 20.6 μg/ml, respectivamente. En el presente estudio se demostró que la CUR inhibe el desenvaine de las L3 a concentraciones menores (< 7 µg/ml) que las de polifenoles mencionados anteriormente.
Se desconoce cuál es el mecanismo por medio del cual la CUR puede inhibir el desenvaine en los tres aislados de H. contortus. Pero es un hecho que la CUR mostró diferencias significativas en su capacidad de inhibir el desenvaine en los tres aislados estudiados. Se ha sugerido que la actividad de bloqueo del desenvaine depende de cómo interactúa la CUR con enzimas, proteínas, ácidos nucleicos, biomoléculas y diferentes tipos de receptores en la vaina de los H. contortus(62). Recientemente, un estudio de metabolómica reveló un panel de metabolitos directamente responsables del desenvaine de las L3, al estar asociado con aminoácidos, así como con las vías del metabolismo de las purinas y el piruvato en H. contortus(63). Es evidente que se deberá investigar el papel de CUR en una o varias de estas vías metabólicas que pueden llevar a inhibir el desenvaine.
Por otro lado, el presente trabajo confirmó que la CUR no afecta la mortalidad de las L3 sin vaina. Dado que la mortalidad causada por diferentes concentraciones de CUR sobre las L3 desenvainadas nunca superó el 10 %. Esto confirma estudios previos que mostraron que el extracto de C. longa ocasiona una mortalidad menor al 10 % cuando se usa a 100 μg/ml(20). Un segundo estudio confirmó la ausencia de algún efecto sobre la mortalidad de L3 de dos extractos de C. longa(53). Todos estos estudios contrastan con el trabajo de Nasai et al(21), quienes proponen que un extracto etanólico de C. longa incrementa la mortalidad de las L3 (78 %) a dosis de 200 mg/ml. Sin embargo, en este último estudio, se usó una gran cantidad de extracto y pudiera no ser viable de replicar en condiciones in vivo. Definitivamente, es importante recalcar que, aun cuando no se logró estimar una CE50 de CUR para la mortalidad de las L3, la prueba de eclosión de huevos demostró que la CUR es capaz de ocasionar cambios que pueden ser compatibles con la inviabilidad de las L1 de H. contortus.
Finalmente, se ha corroborado que los aislados de H. contortus de distintas regiones geográficas presentan diferente susceptibilidad in vitro a los compuestos naturales de las plantas, tanto en la prueba de IEH como la de IDL(64,65). Estos autores sugieren un proceso adaptativo por parte del parásito, para lograr su supervivencia aún en presencia de compuestos secundarios con actividad AH que son ingeridos por el rumiante durante el pastoreo. En este estudio fue posible demostrar que en los diversos bioensayos in vitro los aislados presentaron respuestas diferentes en las concentraciones de CUR evaluadas. Esto fue más evidente para las pruebas de IDL en las que los tres aislados fueron diferentes entre sí. Se desconoce por qué el aislado CENID-SAI, INIFAP presentó la menor sensibilidad al efecto de CUR sobre el desenvaine con respecto a los otros dos aislados de campo de Yucatán.
Conclusiones e implicaciones
La CUR no mostró actividad AH in vitro contra la eclosión de huevos, la migración de L3 o la mortalidad de L3 sin vaina de ninguno de los tres aislados (FMVZ-UADY, Paraíso, CENID-SAI, INIFAP). Sin embargo, se demostró un efecto sobre el desenvaine de las L3, al igual que diferencias entre los tres aislados de H. contortus estudiados.
Conflicto de intereses
Los autores declaran que no existe conflicto de intereses.
Agradecimientos
La presente investigación fue financiada por el CENID SAI, INIFAP a través del proyecto 20454534898 y forma parte del proyecto de Doctorado de Lisandro Encalada Mena en la FMVZ-UADY, México. Así también Lisandro Encalada Mena recibió una beca de estudios de posgrado del programa PRODEP-SEP (UNACAM-122). La producción de huevos y larvas de diferentes aislados de H. contortus fue obtenida del cepario de NGI de la FMVZ-UADY que es un producto del proyecto Cátedras CONAHCYT No. 692.
Literatura citada: