
https://doi.org/10.22319/rmcp.v15i4.6313
Article
Seasonal dynamics of sideoats grama [Bouteloua curtipendula (Mich.) Torr.] in Chihuahua, Mexico: a geostatistical approach
Ireyli Zuluamy Iracheta-Lara a
Carlos Raúl Morales-Nieto b*
Gerardo Méndez-Zamora c
Jaime Neftalí Márquez-Godoy d
Federico Villarreal-Guerrero b
a Universidad Autónoma de Chihuahua. Facultad de Ciencias Agrotecnológicas, Campus 1, Av. Pascual Orozco SN, 31350 Chihuahua, Chihuahua, México.
b Universidad Autónoma de Chihuahua. Facultad de Zootecnia y Ecología, Periférico R. Almada km 1. 31000 Chihuahua, Chihuahua, México.
c Universidad Autónoma de Nuevo León. Facultad de Agronomía. General Escobedo, Nuevo León, México.
d Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). Campo Experimental Valle de Culiacán, Culiacán, Sinaloa, México.
*Corresponding author: cnieto@uach.mx
Abstract:
Phenological data observed on Earth, along with satellite data, are crucial tools for identifying the growing season of vegetation. Using a geostatistical approach, this study aimed to determine the seasonal dynamics of sideoats grama [Bouteloua curtipendula (Michx.) Torr.] in Chihuahua and their relationship with climate variability. The metrics of the start (SOS) and end (EOS) of the growing season of this species in the state of Chihuahua were calculated. In addition, the effect of air temperature and precipitation on the dynamics of SOS and EOS during the 2000-2010 and 2011-2019 periods was evaluated. The treatments considered the three ecological regions (desert, central valleys, and mountain) and the years of recording. The state was studied through three ecological zones: desert (D), central valleys (CV), and sierra (S) for comparison. The SOS and EOS of sideoats grama in each zone were defined annually from Landsat data during the 2000-2019 period based on the dynamics of the Normalized Difference Vegetation Index (NDVI). The SOS ranged from May to June [average Julian day (doy)=174], while the EOS ranged from October to November [average Julian day (doy)=283]. There was a delay in SOS in zone D; the stunted growth of sideoats grama in the spring season may be due to a relative scarcity of water; however, the higher temperature in spring makes it easier to meet the thermal requirements for the species’ growth. These findings suggest that climate variability significantly impacts the seasonal dynamics of sideoats grama, which may influence the management strategies of these ecosystems.
Keywords: Geomatics, Environmental variability, NDVI, Chihuahua.
Received: 12/09/2022
Accepted: 30/07/2024
Introduction
Climate variability, including changes in air temperature and precipitation, significantly impacts the seasonal dynamics of vegetation. Seasonal dynamics refers to the periodic changes in the biological processes of plants throughout the year, influenced by environmental factors(1). Recent studies have shown that variability in rainfall and temperatures can significantly alter plant growth and senescence patterns(2,3). It is highlighted how the increase in global temperature and the decrease in rainfall(4) has led to an extension of the growing season in various regions, highlighting the need to adapt agricultural practices to these new conditions(5,6).
For vegetation monitoring, typical methodologies include recording species-level terrestrial data to perform plant-specific analyses at the local scale(7). At the regional and global levels, satellite data are used to define characteristics and periods of vegetation or landscapes(8,9). Both in situ and satellite phenological data are commonly used to identify the growth phase of vegetation at different scales. Several satellite-derived vegetation indices, such as the normalized difference vegetation index (NDVI) and the enhanced vegetation index (EVI), have been developed to extract phenological parameters from vegetation(10,11). The abovementioned indices focused on analyzing the spatial-temporal differences of phenological phenomena between different biomes or geographical zones(12,13).
In the state of Chihuahua, grasslands, including sideoats grama [Bouteloua curtipendula (Michx.) Torr.], are essential for providing resources for livestock and the ecological stability of the region. Research has shown that changes in patterns, such as flowering and fruiting time, are early signs of the effects of climate variability(14). Conversely, in places where consecutive rainfall events occur sporadically, the increase in temperature may not have significant effects on spring phenology(15,16). Increased rainfall has not shown significant effects on triggering the flowering phase in grasslands(17); in contrast, reduced rainfall induced earlier growth and flowering of herbaceous species in field experiments(18). Therefore, the study of phenology and its relationships with climatic factors would serve to adopt appropriate strategies for grazing activities with a view to the sustainable use of grasslands.
In Chihuahua, rainfall in 2011 was only 156 mm, representing a third of the annual average (i.e., 470 mm), causing huge economic losses. In addition, frost events recorded during the same period in the state were not typical. These events destroyed pastures and caused the death of more than 300,000 head of cattle(19,20). The objectives of this research were to analyze the phenological metrics of the start (SOS) and the end (EOS) of the growing season during 2000-2010 and 2011-2019, which are periods before and after 2011. Additional objectives included determining which climate factor is involved in the dynamics of the SOS and identifying possible trends of change in SOS and EOS during the 2000-2010 and 2011-2019 periods. This will make it possible to assess whether a disturbing phenomenon determines or alters seasonal dynamics and affects grassland growing periods, providing vital information for their sustainable management in the region.
Material and methods
Study area
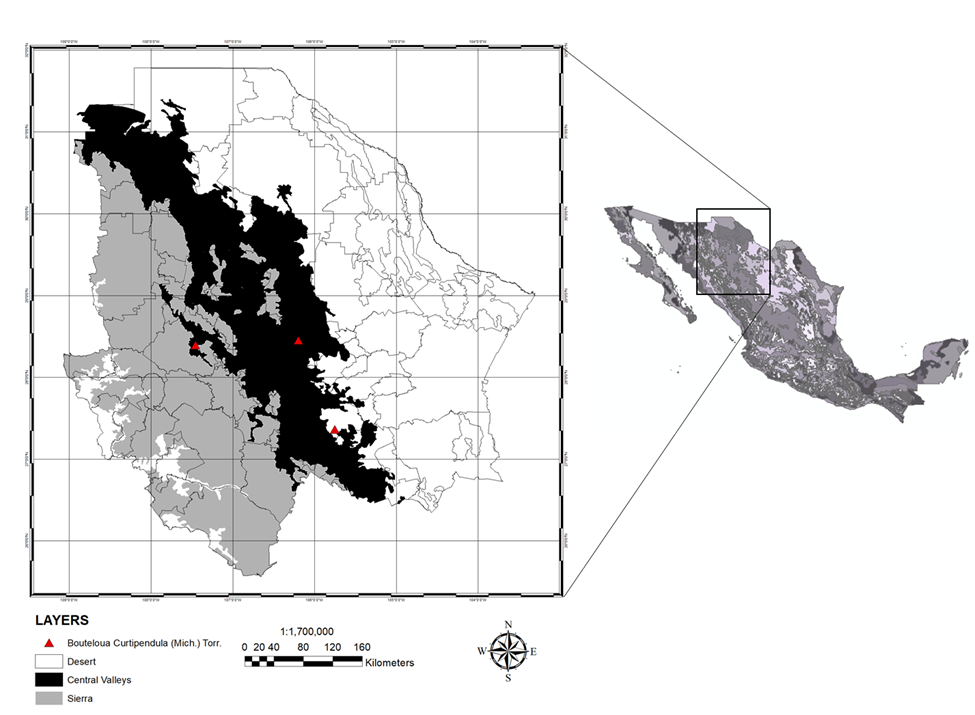
To organize the information, it was decided to classify the state of Chihuahua into ecological zones: Desert (D), Central Valleys (CV), and Sierra (S) according to the climatic conditions they present (Figure 1). Zone D has a dry and semi-warm climate, with daily maximum temperatures of 40 ºC or higher in the summer and daily minimum temperatures of -5 ºC or lower in the winter. On the other hand, the CV zone has a temperate semi-desert to subhumid climate, with daily minimum temperatures of up to -20 ºC in winter, commonly the most extreme minimum temperatures in the state. Annual rainfall ranges between 300 and 550 mm in this region. Finally, the S has a climate with a humid, semi-cold-to-temperate summer and winter with snowfall. Minimum temperatures can drop to -10 °C, while maximum temperatures can reach 30 °C. The climate responds, among other factors, to the altitude above sea level, which on average is 2,400 m, and plains surrounded by peaks 200 to 1,000 m above the plain floor(21).
Figure 1: Study area
Data collection
Precipitation and temperature data were obtained from CONAGUA(22) for the period from 2000 to 2019. On the other hand, the Normalized Difference Vegetation Index (NDVI) was calculated from scenes obtained from the Landsat ETM+7 and Landsat OLI8 (https://glovis.usgs.gov/). In addition, 196 verified records of the presence of sideoats grama [Bouteloua curtipendula (Michx.) Torr.] in each of the three ecological regions into which the state was divided were used.
The calculation of the NDVI (equation 1) was performed on the Google Earth Engine (http://earthengine.google.com/) platform. Scene information was obtained every 16 days for the 2000-2019 period at a spatial resolution of 30 m (WRS Path/Row 32/34 38/42).
NDVI=NIR-RedNIR+Red
Where: NDVI is the Normalized Difference Vegetation Index. NIR and Red are the reflectance values in the near-infrared and red ranges of the electromagnetic spectrum, respectively.
NDVI was obtained from compounds of maximum values, choosing observations with minimal cloud cover and near-nadir views(23). Additional corrections were applied for atmospheric gases, clouds, and aerosols(24). Time series were created with valuable information from NDVI. Although the highest-quality reflectance measurements were selected, likely, the low-quality measurements will still be part of the time series due to cloudy periods. Removing these low-quality values in the NDVI time series was crucial to obtaining a credible phenological metric. There are usually some sequential low points that occur at the beginning and end of the time series, as well as abrupt and extreme low points in the middle of the growing period. First, the issue of the abrupt points [NDVI (t)] corresponding to the middle of the station was addressed; if the difference between NDVI (t) and NDVI (t ± 1) was greater than (NDVImax - NDVImin)/2, then NDVI (t) was replaced by the mean of NDVI (t - 1) and NDVI (t + 1). The entire time series was then smoothed with a 9-point moving average filter. Finally, the low values at the beginning and end of the time series were replaced by the average NDVI value for March, when most NDVI values are similar to the NDVI value of bare soil, due to the limited presence of vegetation on the surface. In this case, NDVI values during the non-growing season are assumed to be constant.
Analysis of phenological function
The typical logistic function retrieves phenological metrics during spring and fall separately; in contrast, the dual logistic function can extract phenological events of spring and fall simultaneously. In this study, equations 2 and 3 were used to obtain SOS and EOS, respectively(25):
SOS=2ln3-2I+S (Equation 2)
EOS=2ln3-2D+E (Equation 3)
Where SOS and EOS denote the day of the year (doy) of the start and end of the season, I and D represent the maximum ascending and descending slopes (inflection points) on the adjusted NDVI curve, respectively, while S and E represent doy when I and D occur on the adjusted NDVI curve.
Statistical analysis
SOS and EOS data were compared at the start and end of the study period rather than calculating the mean multi-pixel trends of SOS and EOS. This was decided to reduce miscalculations. To eliminate the effects of abnormal years on SOS and EOS, the global mean of SOS and EOS during 2000-2010 and 2011-2019 was first obtained separately for each time series. To infer the possible causes of the variations in SOS and EOS, temperature and precipitation records were obtained from the meteorological stations closest to the sampling point(26). The records corresponded to dates before SOS and EOS and the 2000-2010 and 2011-2019 periods.
SPD and EPD data were compared for their differences between zones D, CV, and S with a univariate analysis of variance (ANOVA). This was done through a 3x2 factorial arrangement, three regions: D, CV, and S, and two periods: 2000 to 2010 and 2011 to 2019. Of the 196 records of the presence of grasslands, a coordinate of each ecological zone was randomly selected in order to compare the vegetation in different areas and how it responds to changes in climatic conditions (Figure 1). In addition, a correlation analysis was performed to evaluate the relationship between SPD and EPD with the climatic variables of each region and period.
Results and discussion
Environmental conditions
The environmental conditions average (precipitation and temperature) during the 2000-2019 period showed that the spring temperature in zone D, where the record of the sideoats grama was analyzed, was higher than in zones CV and S. Meanwhile, the precipitation of the spring season was higher in zone D than in zones CV and S.
As reported in previous studies, spring phenological events are particularly sensitive to temperature, and the warming experienced in recent decades has already shown effects on phenology, triggering earlier spring phenological events and extending the growing period(27). Zone D was also warmer and drier in autumn than zones CV and S. In all three ecological zones, autumn is cooler and wetter than spring (Table 1). Authors stated(28) that temperature is the main climatic factor affecting plant phenology. They also reported that increased air temperature affects phenology and can be easily detected in phenological data.
Table 1: Environmental conditions in the Sierra, Central Valleys, and Desert
ST= spring temperature; SP= spring precipitation; AT= autumn temperature; AP= autumn precipitation.
General comparison of SOS and EOS between ecological zones
The averages for SPD and EPD were on the 174th and 283rd days of the year, respectively; locating these days in months, they would be May-June and October. In zone D, SPD and EPD occurred on the 172nd and 265th d of the year, respectively, earlier (P<0.05) in the year compared to the other two zones (doy 180 and 286). The SOS occurred earlier in zone D than in zones CV and S. Both SOS and EOS occurred significantly (P<0.05) earlier in zone D (doy 172 and 265, respectively) than in zones CV (doy 180 and 286, respectively) and D (doy 179 and 297, respectively). Regarding the annual fluctuations of SOS and EOS, the mean standard deviation (SD) of EOS (25.47 d) was greater than that of SOS (18.53 d). The highest SDs of SOS and EOS corresponded to zone D (21.6 and 25.15 d, respectively) compared to zones CV (19.5 and 25.05 d, respectively) and S (11.5 and 12.41 d, respectively). Phenological models based on satellite data indicate water availability as a determining environmental condition in SPD in North American meadows(7); for its part, temperature was a determining factor in triggering spring phenological events in woody plants of temperate ecosystems(29) and herbaceous plants of alpine ecosystems(30).
SOS and EOS response rates to temperature and precipitation
Although there was a 0.33-d delay for SOS in zone D during 2000-2019, SOS occurred 0.20 and 0.09 d earlier for zones CV and S, respectively (Figure 2a). Meanwhile, the decrease in spring precipitation, which happened before the start of the season, caused an SOS delay of 0.42, 0.24, and 0.32 d mm-1 for zones D, CV, and S, respectively (Figure 2b). For zone D, the sensitivity of SOS to the preceding temperature and precipitation, which were recorded before the start of the season, was significantly higher than in zones CV and S. In autumn, warmer temperatures delayed SOS at rates of 0.94 and 0.35 d ºC-1 for zones CV and S, respectively, while extending SOS by 0.72 d ºC-1 for zone D (Figure 2c).
Figure 2: SOS response to preceding temperature (a) and precipitation (c), and SOS response rates due to preceding temperature (b) and precipitation (d) at EOS
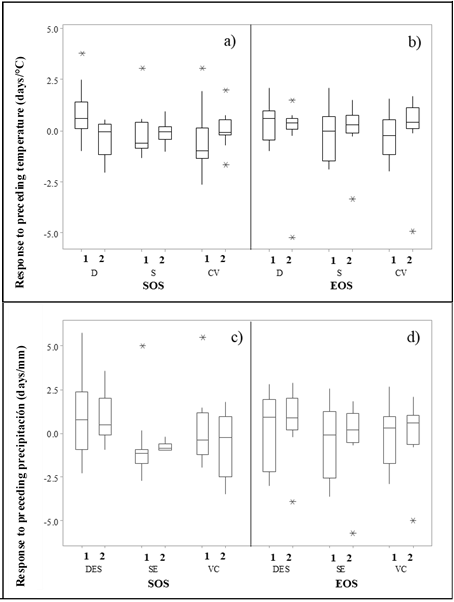
1, period from 2000 to 2010; 2, period from 2011 to 2019. D= desert; S= sierra; CV= central valleys.
Some authors(31) reported that, during 1959-1996, the spring phenological events of a large number of species were 6.3 days early on average, while those of autumn experienced a delay of 4.5 days on average; thus, the growing season extended by 10.8 days on average(32,33). The reduction in rainfall caused a delay of 0.89, 0.02, and 0.45 d mm-1 in EOS for zones D, CV, and S, respectively (Figure 2d). Finally, zone D showed a significantly lower sensitivity to the preceding temperature but a significantly higher sensitivity to the preceding precipitation than zones CV and S. Anticipated phenological patterns could alter the distribution of sideoats grama plant resources, have implications for pollination systems, and increase the size, species richness, and intraspecific genetic diversity of the soil seed bank(34).
SOS and EOS change trends
To report overall SOS and EOS changes in the ecological zones studied, the annual mean SOS and EOS between 2000-2010 and 2011-2019 were compared. During 2010-2019, SOS and EOS were delayed by 16 (P<0.05) and 18 (P<0.05) days, respectively, compared to 2000-2019 (Figure 3).
The sideoats grama results showed that the mean 2011-2019 SOS was 16 d later (P<0.05) than the 2000-2010 SOS mean for zone D (Figure 3). On the other hand, SOS was 42 d early for zone CV (P<0.05) and 3 d early for zone S (P<0.05). Meanwhile, EOS showed no significant changes (P>0.05) when comparing the two periods for zones CV and S. Nonetheless, a significant lag was identified for both SOS and EOS from 2000-2010 to 2011-2019 (P<0.05) in zone D. During 2011-2019, SOS and EOS were 6 (P<0.05) and 18 (P<0.05) days late, respectively, compared to 2000-2010.
Figure 3: General changes in SOS and EOS in three ecological zones affecting the species. 1, period from 2000 to 2010; 2, period from 2011 to 2019.
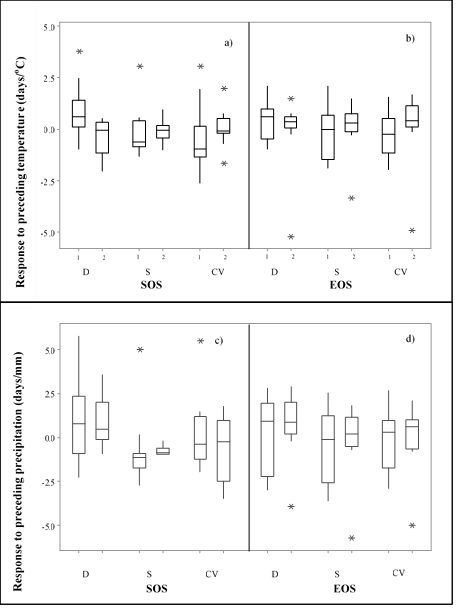
1, period from 2000 to 2010; 2, period from 2011 to 2019. D= desert; S= sierra; CV= central valleys.
This study found significant negative correlations (P<0.05) between SOS and preceding precipitation and temperature. This means that higher temperatures and rainfall recorded before the start of the season can bring forward the start of the growing season (SOS). Nevertheless, the zones showed significant correlations (P<0.05) between SOS and the preceding precipitation than between EOS and the antecedent temperature. Therefore, recorded precipitation rather than temperature may primarily control the start of the growing season in most of the study area. This is consistent with the results of previous studies based on data from greening dates, derived from satellites, and terrestrial meteorological data(35). On the other hand, some experimental and phenology modelling studies have highlighted the critical effects of water availability on SOS in the North American meadows(7,36) and Mongolian meadows(2,11). Temperature has been reported to be a key factor in triggering spring phenological events of plants in temperate ecosystems(37) and herbaceous species in alpine ecosystems(38). The predominant role of temperature in SOS has also been observed in grasslands in the Middle East and in the Tibetan Plateau(39,40).
Likewise, the ecological zones of the study area showed significant positive correlations (P<0.05) between EOS and the preceding temperature, indicating that temperature may be the most critical factor in regulating seasonal vegetation dynamics in grasslands. Although temperature was the dominant factor, precipitation was also positively correlated with EOS. Therefore, warmer and wetter autumn conditions could lead to a delay in the process of leaf senescence.
Behavior of SOS and EOS between ecological zones during 2000-2010 and 2011-2019
Factor analysis revealed that the average day of the year (doy) for the start of the growing season (SOS) of the species was 160.55 (P<0.05). Significant effects in the factorial design depended on the zone (D, CV, S) for both SOS and the end of the growing season (EOS). The factor that caused the biggest change in SOS and EOS (i.e., variability in doy) was the ecological zone in which the vegetation grows. The interaction between the ecological zones and the periods analyzed significantly influenced both SOS and EOS (P<0.005).
Conclusions and implications
This research has provided a detailed understanding of the seasonal dynamics of sideoats grama [Bouteloua curtipendula (Michx.) Torr.] in different ecological zones of the state of Chihuahua, Mexico, using a geostatistical approach supported by satellite data and in situ phenological observations. The results show that climate variability, particularly in temperature and precipitation, affects the start (SOS) and end (EOS) of its growing season. It was observed that in the desert zone (D), SOS occurs earlier due to higher temperatures. In addition, decreased precipitation delayed SOS in all zones, while in autumn, warmer temperatures extended EOS in zone D. Understanding the seasonal dynamics of this species is essential to develop sustainable management practices that ensure the resilience of these ecosystems to climate change. The results obtained provide a robust scientific basis for informed decision-making in the management of natural resources in the region, promoting sustainability and adaptation to new environmental conditions.
Conflict of interest
The authors declare that they have no conflict of interest.
Literature cited:
- IPCC. Climate change 2014: Impacts, adaptation, and vulnerability. Part A: Global and sectoral aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge Univ Press; 2014.
- Smith J, Doe J, Clark H. Temporal and spatial variability in phenological responses of grass species. J Ecol 2020;108(5):1234-1245.
- Liu Q, Fu YH, Zeng Z, Huang M, Li X, Piao S. Temperature, precipitation, and insolation effects on autumn vegetation phenology in temperate China. Glob Change Biol 2019;25(3):644-655.
- Sánchez, DH, Navarro-Gómez CJ, Rentería M, Rose JF, Sánchez-Navarro JR. Evolution of the groundwater system in the Chihuahua-Sacramento aquifer due to climatic and anthropogenic factors. J Water Clim Change 2022;13(2):645-663.
- Jones MB, Donnelly A. Responses of Irish vegetation to future climate change. Biol Environ Proc R Ir Acad 2018;118(2):157-168.
- Williams K, Clark, J. Global warming extends the growing season across ecosystems. Environ Res Lett 2021;16(1):14-23.
- Schwartz MD. Phenology: An integrative environmental science. Berlin, Germany Springer; 2013:170-171.
- Badeck FW, Bondeau AK, Böttcher D, Doktor W, Lucht J, Schaber S. Responses of spring phenology to climate change. New Phytol 2004;162:295-309.
- Richardson AD, Keenan TF, Migliavacca M, Ryu Y, Sonnentag O, Toomey M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric For Meteorol 2017;169:156-173.
- Crimmins TM, Crimmins MA, Bertelsen C. Complex responses to climate drivers in onset of spring flowering across a semi-arid elevation gradient. J Ecol 2010;98:042-1051.
- Lesica P, Kittelson P. Precipitation and temperature are associated with advanced flowering phenology in a semi-arid grassland. J Arid Environ 2010;74:1013-1017.
- Zhang X, Friedl MA, Schaaf CB, Strahler AH, Hodges JCF, Gao F, Reed BC, Huete A. Monitoring vegetation phenology using modis. Remote Sens Environ 2003;84:471-475.
- Delbart N, Picard G, Le Toan T, Kergoat K, Quegan S, Woodward I, Dye D, Fedotova V. Spring phenology in boreal Eurasia over a nearly century time scale. Glob Change Biol 2008;14:603-614.
- Stumpf F, Schneider MK, Keller A, Rentschler T, Meuli RG, Liebisch, F. Spatial monitoring of grassland management using multi-temporal satellite imagery. Ecol Indic 2020;113:106-121.
- Schwartz MD. Phenology: An integrative environmental science. Springer 2013;1:20.
- Piao S, Tan J, Chen A, Fu YH, Ciais P, Liu Q, Janssens IA, Vicca S, Zeng Z, Jeong SJ. Leaf onset in the Northern Hemisphere triggered by daytime temperature. Nat Commun 2015;6:69-81.
- Bloor JM, Pichon P, Falcimagne R, Leadley P, Soussana JF. Effects of warming, summer drought, and CO2 enrichment on aboveground biomass production, flowering phenology, and community structure in an upland grassland ecosystem. Ecosystems 2010;13:888-900.
- Hovenden MJ, Wills KE, Vander-Schoor JK, Williams AL, Newton PC. Flowering phenology in a species-rich temperate grassland is sensitive to warming but not elevated CO2. New Phytol 2008;178:815-822.
- Zelikova TJ, Williams DG, Hoenigman R, Blumenthal DM, Morgan JA, Pendall E. Seasonality of soil moisture mediates responses of ecosystem phenology to elevated CO2 and warming in a semi-arid grassland. J Ecol 2015;103:1119-1130.
- Piazza V. Estimación satelital de la productividad primaria neta aérea de la vegetación herbácea del Caldenal [disertación doctoral]. Buenos Aires, Argentina; 2006.
- Morales-Nieto CR, Quero-Carrillo A, Pérez-Pérez J, Hernández-Garay A, Le-Blan O. Caracterización morfológica de poblaciones nativas de pasto banderita [(Michx.) Torr.] en México. Agrociencia 2008;42(7):767-775.
- Breman H, De Wit C. Rangeland productivity and exploitation in the Sahel. Science 1983;221:1341–1347.
- Butt B, Turner MD, Singh A, Brottem L. Use of modis NDVI to evaluate changing latitudinal gradients of rangeland phenology in sudano-sahelian West Africa. Remote Sens Environ 2011;115:3367-3376.
- Arzani H, Zohdi M, Fish E, Zahedi-Amiri G, Nikkhah A, Wester D. Phenological effects on forage quality of five grass species. J Range Management 2004;57:624–629.
- Gorlier A, Lonati M, Renna M, Lussiana C, Lombardi G, Battaglini L. Changes in pasture and cow milk compositions during a summer transhumance in the Western Italian Alps. J Appl Bot Food Qual 2013;(85):216-231.
- Ortega-Gaucin D, Velasco I. Aspectos socioeconómicos y ambientales de las sequías en México. Aqua-LAC 2013;5(2):78-90.
- Gonsamo A, Chen JM. Continuous observation of leaf area index at fluxnet-Canada sites. Agric For Meteorol 2014;189:168-174.
- INEGI. Instituto Nacional de Estadística, Geografía e Informática 2003. Síntesis de Información Geográfica del Estado de Chihuahua, 1a.ed. Editorial INEGI. México. 2013.
- CONAGUA. Comisión Nacional del Agua. Capacidad de Almacenamiento y Uso de las Principales Presas de México. http://www.conagua.gob.mx/atlas/atlas.html?seccion=2mapa=5. Consultado 4 Abr, 2021.
- Chen X, Li J, Xu L, Liu L, Ding D. Modeling green up date of dominant grass species in the inner Mongolian grassland using air temperature and precipitation data. Int J Biometeorol 2014;58:463-471.
- Vermote E. Vermeulen. Atmospheric correction algorithm: Spectral reflectances (MOD09), ATBD version 4; Department of Geography, University of Maryland: College Park, MD, USA. 1999.
- Busetto L, Colombo R, Migliavacca M, Cremonese E, Meroni M, Galvagno M, et al. Remote sensing of larch phenological cycle and analysis of relationships with climate in the Alpine region. Glob Change Biol 2010;16:2504-2517.
- Menzel A. Trends in phenological phases in Europe between 1951 and 1996. Int J Biometeorol 2000;44:76–81.
- Peñuelas J, Filella I. Responses to a warming world. Science 2001;294:793–795.
- Chen X, Xu L. Phenological responses of Ulmus pumila (Siberian Elm) to climate change in the temperate zone of China. Int J Biometeorol 2012;56:695-706.
- Chen X, An S, Inouye MD. Temperature and snowfall trigger alpine vegetation green-up on the world’s roof. Glob Change Biol 2015;21:3635-3646.
- Peñuelas J, Filella I. Responses to a warming world. Science 2001;294(3):793-795.
- Roetzer T, Wittenzeller M, Haeckel H, Nekovar J. Phenology in central Europe–differences and trends of spring phenophases in urban and rural areas. Int J Biometeorol 2000;44:60–66.
- Walkovszky A. Changes in phenology of the locust tree (Robinia pseudoacacia L.) in Hungary. Int J Biometeorol 1998;41:155–160.