https://doi.org/10.22319/rmcp.v15i4.6313
Artículo
Dinámica estacional de pasto banderita [Bouteloua curtipendula (Mich.) Torr.] en Chihuahua, México: enfoque geoestadístico
Ireyli Zuluamy Iracheta-Lara a
Carlos Raúl Morales-Nieto b*
Gerardo Méndez-Zamora c
Jaime Neftalí Márquez-Godoy d
Federico Villarreal-Guerrero b
a Universidad Autónoma de Chihuahua. Facultad de Ciencias Agrotecnológicas, Campus 1, Av. Pascual Orozco SN, 31350 Chihuahua, Chihuahua, México.
b Universidad Autónoma de Chihuahua. Facultad de Zootecnia y Ecología, Periférico R. Almada km 1. 31000 Chihuahua, Chihuahua, México.
c Universidad Autónoma de Nuevo León. Facultad de Agronomía. General Escobedo, Nuevo León, México.
d Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). Campo Experimental Valle de Culiacán, Culiacán, Sinaloa, México.
*Autor de correspondencia: cnieto@uach.mx
Resumen:
Los datos fenológicos observados en tierra, junto con datos satelitales, son herramientas cruciales para identificar la estación de crecimiento de la vegetación. Utilizando un enfoque geoestadístico, este estudio tuvo como objetivo determinar la dinámica estacional del pasto banderita [Bouteloua curtipendula (Michx.) Torr.] en Chihuahua y su relación con la variabilidad climática. Se calcularon las métricas del inicio (SOS) y del final (EOS) de la temporada de crecimiento de esta especie en el estado de Chihuahua. Además, se evaluó el efecto de la temperatura del aire y la precipitación en la dinámica del SOS y el EOS durante los periodos 2000-2010 y 2011-2019. Los tratamientos consideraron las tres regiones ecológicas (desierto, valles centrales y sierra) y los años de registro. El estado se estudió a través de tres zonas ecológicas: desierto (D), valles centrales (CV) y sierra (S) para su comparación. El SOS y el EOS del pasto banderita en cada zona se definieron anualmente a partir de datos Landsat durante el periodo 2000-2019, basándose en la dinámica del Índice de Vegetación de Diferencia Normalizada (NDVI). El SOS osciló entre mayo y junio (promedio de día juliano [doy]=174), mientras que el EOS osciló entre octubre y noviembre (promedio de día juliano [doy]=283). Se observó un retraso en el SOS en la zona D; el retraso en el crecimiento del pasto banderita en la estación primaveral puede deberse a una relativa escasez de agua, aunque la mayor temperatura en primavera facilita el cumplimiento de los requisitos térmicos para el crecimiento de la especie. Estos hallazgos sugieren que la variabilidad climática tiene un impacto significativo en la dinámica estacional del pasto banderita, lo que puede influir en las estrategias de manejo de estos ecosistemas.
Palabras clave: Geomática, Variabilidad ambiental, NDVI, Chihuahua.
Recibido: 12/09/2022
Aceptado: 30/07/2024
Introducción
La variabilidad climática, incluyendo cambios en la temperatura del aire y las precipitaciones, tiene un impacto significativo en la dinámica estacional de la vegetación. La dinámica estacional se refiere a los cambios periódicos en los procesos biológicos de las plantas a lo largo del año, influenciados por factores ambientales(1). Estudios recientes han demostrado que la variabilidad en las precipitaciones y las temperaturas pueden alterar significativamente los patrones de crecimiento y senescencia de las plantas(2,3). Se subraya cómo el aumento de la temperatura global y la disminución de las precipitaciones(4) ha llevado a una extensión de la temporada de crecimiento en diversas regiones, destacando la necesidad de adaptar las prácticas agrícolas a estas nuevas condiciones(5,6).
Para el seguimiento de la vegetación, las metodologías habituales incluyen el registro de datos terrestres a nivel de especie para realizar análisis específicos de las plantas a escala local(7). A nivel regional y global, los datos satelitales se utilizan para definir características y periodos de la vegetación o de los paisajes(8,9). Tanto los datos fenológicos in situ como los obtenidos por satélite se utilizan habitualmente para identificar la fase de crecimiento de la vegetación a diferentes escalas. Se han desarrollado varios índices de vegetación derivados de satélites, como el índice de vegetación de diferencia normalizada (NDVI) y el índice de vegetación mejorado (EVI), para extraer parámetros fenológicos de la vegetación(10,11). El uso de los índices mencionados se ha centrado en el análisis de las diferencias espacio-temporales de los fenómenos fenológicos entre diversos biomas o zonas geográficas(12,13).
En el estado de Chihuahua, los pastizales, incluido el pasto banderita [Bouteloua curtipendula (Michx.) Torr.], son fundamentales para la provisión de recursos para la ganadería y la estabilidad ecológica de la región. Investigaciones han demostrado que los cambios en los patrones, como el tiempo de floración y fructificación, son señales tempranas de los efectos de la variabilidad climática(14). Por el contrario, en lugares donde los eventos de lluvia consecutivos ocurren esporádicamente, el aumento de la temperatura puede no tener efectos significativos en la fenología primaveral(15,16). El aumento de las precipitaciones no ha mostrado efectos significativos en el desencadenamiento de la fase de floración en pastizales(17), mientras que la reducción de las precipitaciones indujo un crecimiento y una floración más tempranos de las especies herbáceas en experimentos de campo(18). Por lo tanto, el estudio de la fenología y sus relaciones con los factores climáticos serviría para adoptar estrategias adecuadas para las actividades de pastoreo, con vistas al uso sostenible de los pastizales.
En Chihuahua, la precipitación durante el 2011 fue de sólo 156 mm, lo que representa un tercio del promedio anual (es decir, 470 mm), causando enormes pérdidas económicas. Además, los eventos de heladas registrados durante el mismo periodo en el estado no fueron típicos. Estos eventos destruyeron los pastos y causaron la muerte de más de 300,000 bovinos(19,20). Los objetivos de esta investigación fueron analizar las métricas fenológicas del inicio (SOS) y del final (EOS) de la temporada de crecimiento durante 2000-2010 y 2011-2019, que son períodos anteriores y posteriores a 2011. Los objetivos adicionales incluyeron determinar qué factor climático está involucrado en la dinámica del SOS e identificar posibles tendencias de cambio en el SOS, así como en el EOS, durante los períodos de 2000-2010 y 2011-2019. Esto permitirá evaluar si un fenómeno perturbador determina o altera las dinámicas estacionales e incide en los períodos de crecimiento de los pastizales, proporcionando información vital para su gestión sostenible en la región.
Material y métodos
Área de estudio
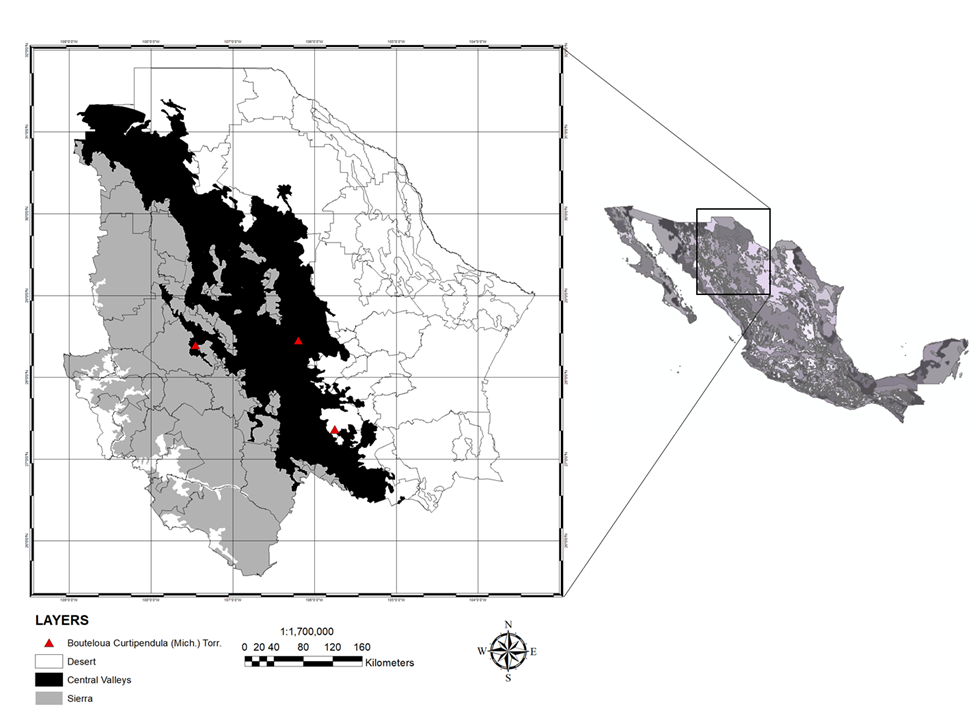
Para la organización de la información se decidió clasificar el estado de Chihuahua en zonas ecológicas: Desierto (D), Valles Centrales (CV) y Sierra (S) de acuerdo a las condiciones climáticas que presentan (Figura 1). La zona D tiene un clima seco y semicálido, con temperaturas máximas diarias de 40 ºC o superiores en el verano y temperaturas mínimas diarias de -5 ºC o menores en invierno. Por otra parte, la zona CV, tiene un clima templado de semidesértico a subhúmedo, en invierno con temperaturas mínimas diarias de hasta -20 ºC, comúnmente las temperaturas mínimas más extremas del estado. La precipitación anual oscila entre 300 y 550 mm en esta zona. Finalmente, la S tiene un clima con verano húmedo de semifrío a templado e invierno con nevadas. Las temperaturas mínimas pueden descender hasta -10 °C, mientras que las máximas pueden alcanzar los 30 °C. El clima responde entre otros factores a la altura sobre el nivel del mar que en promedio está a 2,400 m, llanuras rodeadas por picos que con 200 a 1,000 m por arriba del piso de la llanura(21).
Figura 1: Área de estudio
Recolección de datos
Los datos de precipitación y temperatura se obtuvieron de CONAGUA(22) para el periodo del 2000 al 2019. Por otra parte, el Índice de Vegetación de Diferencia Normalizada (NDVI, por sus siglas en inglés) se calculó a partir de escenas obtenidas del Landsat ETM+7 y Landsat OLI8 (https://glovis.usgs.gov/). Además, 196 registros verificados de la presencia del pasto banderita [Bouteluoa curtipendula (Michx.) Torr.] en cada una de las tres regiones ecológicas en que se dividió el estado.
El cálculo del NDVI (ecuación 1) se realizó en la plataforma Google Earth Engine (http://earthengine.google.com/). Se obtuvo información de escenas de cada 16 días para el periodo 2000-2019 a una resolución espacial de 30 m (WRS Path/Row 32/34 38/42).
NDVI=NIR-RedNIR+Red
Donde, NDVI es el Índice de Vegetación de Diferencia Normalizada. NIR y Red son los valores de reflectancia en los rangos infrarrojo cercano y rojo del espectro electromagnético, respectivamente.
El NDVI se obtuvo a partir de compuestos de valores máximos, eligiendo observaciones con mínima cobertura de nubes y vistas cercanas al nadir(23). Se aplicaron correcciones adicionales para gases atmosféricos, nubes y aerosoles(24). Se elaboraron series temporales con información valiosa del NDVI. Aunque se seleccionaron las mediciones de reflectancia de mayor calidad, es probable que las mediciones de baja calidad sigan formando parte de las series temporales debido a los períodos nublados. La eliminación de estos valores de baja calidad en las series temporales de NDVI fue de crucial importancia para obtener una métrica fenológica creíble. Por lo general, hay algunos puntos bajos secuenciales que se producen al principio y al final de la serie temporal, así como puntos bajos abruptos y extremos en la mitad del período de crecimiento. En primer lugar, se abordó la cuestión de los puntos abruptos [NDVI (t)] correspondientes a la mitad de la estación; si la diferencia entre el NDVI (t) y el NDVI (t ± 1) era mayor que (NDVImax - NDVImin)/2, entonces el NDVI (t) se sustituyó por la media del NDVI (t - 1) y el NDVI (t+1). A continuación, se suavizó toda la serie temporal con un filtro de media móvil de 9 puntos. Por último, los valores bajos al principio y al final de la serie temporal se sustituyeron por el valor medio del NDVI de marzo, cuando la mayoría de los valores del NDVI son similares al valor del NDVI del suelo desnudo, debido a la limitada presencia de vegetación en la superficie. En este caso, se asumió que los valores de NDVI durante la temporada de no crecimiento son constantes.
Análisis de la función fenológica
La función logística típica recupera la métrica fenológica durante la primavera y el otoño por separado, mientras que la función logística dual puede extraer los eventos fenológicos de primavera y otoño, simultáneamente. En este estudio, se utilizaron las ecuaciones 2 y 3 para obtener el SOS y el EOS, respectivamente(25):
SOS=2ln3-2I+S (Ecuación 2)
EOS=2ln3-2D+E (Ecuación 3)
Donde, SOS y EOS denotan el día del año (doy) del inicio y del final de la estación, I y D representan las máximas pendientes ascendentes y descendentes (puntos de inflexión) en la curva NDVI ajustada, respectivamente, mientras que S y E representan el doy cuando I y D se producen en la curva NDVI ajustada.
Análisis estadístico
Los datos de SOS y EOS se compararon al inicio y al final del periodo de estudio, en lugar de calcular las tendencias medias de múltiples píxeles de SOS y EOS. Esto se decidió para reducir los errores de cálculo. Para eliminar los efectos de los años anormales en SOS y EOS, se obtuvo primero la media global de SOS y EOS durante 2000-2010 y 2011-2019, por separado para cada serie temporal. Para inferir sobre las posibles causas de las variaciones del SOS y EOS, se obtuvieron registros de temperatura y precipitación de las estaciones meteorológicas más cercanas al punto de muestreo(26). Los registros correspondieron a fechas anteriores al SOS y EOS y del periodo 2000-2010 y 2011-2019.
Los datos de SPD y EPD se compararon sus diferencias entre las zonas D, CV y S con un análisis de varianza univariante (ANOVA). Esto se realizó a través de un arreglo factorial 3x2, tres regiones: D, CV y S y dos periodos 2000 a 2010 y 2011 a 2019. De los 196 registros de presencia de pastizales, se seleccionó al azar una coordenada de cada zona ecológica con el fin de realizar una comparación de la vegetación en diferentes áreas y como ésta responde a cambios en las condiciones climáticas (Figura 1). Además, se realizó un análisis de correlación para evaluar la relación entre SPD y EPD con las variables climáticas de cada región y periodo.
Resultados y discusión
Condiciones ambientales
Las condiciones ambientales medias (precipitación y temperatura) durante el periodo 2000-2019 mostraron que la temperatura primaveral en la zona D donde se analizó el registro del pasto banderita fue mayor que en las zonas CV y S. Mientras tanto, la precipitación de la estación primaveral fue mayor en la zona D que en las zonas CV y S.
Como se ha informado en estudios anteriores, los eventos fenológicos primaverales son particularmente sensibles a la temperatura, y el calentamiento experimentado en las últimas décadas ya ha mostrado efectos sobre la fenología, desencadenando eventos fenológicos primaverales más tempranos y extendiendo el periodo de crecimiento(27). En otoño, la zona D también fue más cálida y seca que las zonas CV y S. En las tres zonas ecológicas, el otoño es más fresco y húmedo que la primavera (Cuadro 1). Autores afirmaron(28) que el principal factor climático que afecta a la fenología de las plantas es la temperatura. También informaron de que el aumento de la temperatura del aire afecta a la fenología y puede detectarse fácilmente en los datos fenológicos.
Cuadro 1: Condiciones medioambientales en la Sierra, Valles Centrales y Desierto
ST= temperatura de primavera; SP= precipitación de primavera; AT= temperatura de otoño; AP= precipitación de otoño.
Comparación general del SOS y el EOS entre zonas ecológicas
Las medias para SPD y EPD fueron en los días 174 y 283 del año, respectivamente, ubicando estos días en meses serían mayo-junio y octubre. Procurar una redacción más sucinta por ejemplo en la zona D, SPD y EPD ocurrieron en los días 172 y 265 del año, respectivamente, más temprano (P<0.05) en el año en comparación a las otras dos zonas (doy 180 y 286). El SOS se produjo antes en la zona D que en las zonas CV y S. Tanto el SOS como el EOS se produjeron significativamente (P<0,05) antes en la zona D (doy 172 y 265, respectivamente) que en las zonas CV (doy 180 y 286, respectivamente) y D (doy 179 y 297, respectivamente). En cuanto a las fluctuaciones anuales del SOS y el EOS, la desviación estándar (DE) media del EOS (25.47 días) fue mayor que la del SOS (18.53 días). Las mayores DE de SOS y EOS correspondieron a la zona D (21.6 y 25.15 días, respectivamente), en comparación con las zonas CV (19.5 y 25.05 días, respectivamente) y S (11.5 y 12.41 días, respectivamente). Los modelos fenológicos basados en datos satelitales indican la disponibilidad de agua como una condición ambiental determinante en el SPD en praderas norteamericanas(7); mientras que la temperatura fue determinante para desencadenar eventos fenológicos primaverales en leñosas de ecosistemas templados(29) y herbáceas de los ecosistemas alpinos(30).
Tasas de respuesta del SOS y el EOS a la temperatura y la precipitación
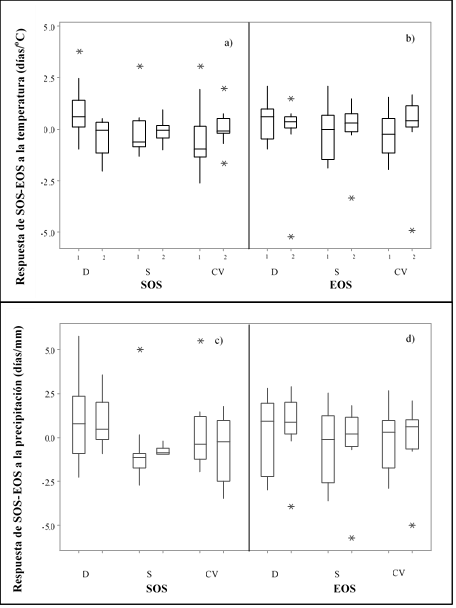
Aunque se produjo un retraso de 0.33 días para el SOS en la zona D durante 2000-2019, el SOS se produjo 0.20 y 0.09 días antes para las zonas CV y S, respectivamente (Figura 2a). Mientras tanto, la disminución de la precipitación primaveral, que se produjo antes del inicio de la estación, provocó un retraso del SOS de 0.42, 0.24 y 0.32 días mm-1 para las zonas D, CV y S, respectivamente (Figura 2b). Para la zona D, la sensibilidad del SOS a la temperatura y la precipitación precedente, que se registraron antes del inicio de la estación, fue significativamente mayor que en las zonas CV y S. En otoño, las temperaturas más cálidas retrasaron el SOS a tasas de 0.94 y 0.35 días ºC-1 para las zonas CV y S, respectivamente, mientras que extendieron el SOS en 0.72 días ºC-1 para la zona D (Figura 2c).
Figura 2: Respuesta del SOS a la temperatura antecedente (a) y a la precipitación (c), y tasas de respuesta del SOS debido a la temperatura antecedente (b) y a la precipitación (d) al EOS
1, periodo de 2000-2010; 2, periodo de 2011-2019. D= desierto; S= sierra; CV= valles centrales.
Algunos autores(31) reportaron que durante 1959-1996, los eventos fenológicos de primavera de un gran número de especies se adelantaron en promedio 6.3 días, mientras que los de otoño han experimentado un retraso de 4.5 días en promedio; así, la estación de crecimiento se extendió en 10.8 días en promedio(32,33). La reducción de las precipitaciones provocó un retraso en el EOS de 0.89, 0.02 y 0.45 días mm-1 para las zonas D, CV y S, respectivamente (Figura 2d). Finalmente, la zona D mostró una sensibilidad significativamente menor a la temperatura antecedente, pero una sensibilidad significativamente mayor a la precipitación antecedente que las zonas CV y S. Los patrones fenológicos anticipados pueden alterar la distribución de los recursos vegetales del pasto banderita, tener implicaciones para los sistemas de polinización y aumentar el tamaño, la riqueza de especies y la diversidad genética intraespecífica del banco de semillas del suelo(34).
Tendencias de cambio de SOS y EOS
Para informar de los cambios generales de SOS y EOS en las zonas ecológicas estudiadas, se comparó la media anual de SOS y EOS entre 2000-2010 y 2011-2019. Durante 2010-2019, el SOS y el EOS se retrasaron 16 (P<0.05) y 18 (P<0.05) días, respectivamente, en comparación con 2000-2019 (Figura 3).
Los resultados del pasto banderita mostraron que la media del SOS de 2011-2019 fue 16 días más tardía (P<0.05) que la media del SOS de 2000-2010 para la zona D (Figura 3). Por el contrario, el SOS se adelantó 42 días para la zona CV (P<0.05) y 3 días para la zona S (P<0.05). Mientras tanto, el EOS no mostró cambios significativos (P>0.05) al comparar los dos períodos para las zonas CV y S. Sin embargo, se identificó un retraso significativo tanto para SOS como para EOS desde 2000-2010 hasta 2011-2019 (P<0.05) en la zona D. Durante 2011-2019, el SOS y el EOS se retrasaron 16 (P<0.05) y 18 (P<0.05) días respectivamente en comparación con 2000-2010.
Figura 3: Cambios generales en SOS y EOS en tres zonas ecológicas en afectación a la especie
1, periodo de 2000-2010; 2, periodo de 2011-2019. D= desierto; S= sierra; CV= valles centrales.
Este estudio encontró correlaciones negativas significativas (P<0.05) entre el SOS y la precipitación y la temperatura precedente. Esto significa que temperaturas y precipitaciones más elevadas registradas antes del inicio de la estación pueden adelantar el inicio de la estación de crecimiento (SOS). Sin embargo, las zonas mostraron correlaciones significativas (P<0,05) entre el SOS y la precipitación antecedente que entre el EOS y la temperatura antecedente. Por lo tanto, la precipitación registrada en lugar de la temperatura puede controlar principalmente el inicio de la temporada de crecimiento en la mayor parte del área de estudio. Esto concuerda con los resultados de estudios anteriores basados en datos de fechas de verdeo, derivados de satélites y datos meteorológicos terrestres(35). Por otro lado, algunos estudios experimentales y de modelización de la fenología han destacado los efectos críticos de la disponibilidad de agua en la SOS en las praderas norteamericanas(7,36) y mongolas(2,11). Se ha informado que la temperatura es un factor clave para desencadenar los eventos fenológicos primaverales de las plantas en los ecosistemas templados(37) y de las especies herbáceas en los ecosistemas alpinos(38). El papel predominante de la temperatura en el SOS también se ha observado en los pastizales de Oriente Medio y en la meseta Tibetana(39).
Asimismo, las zonas ecológicas del área de estudio mostraron correlaciones positivas significativas (P<0.05) entre el EOS y la temperatura antecedente, lo que indica que la temperatura puede ser el factor más importante en la regulación de la dinámica estacional de la vegetación en los pastizales. Aunque la temperatura fue el factor dominante, la precipitación también se correlacionó positivamente con el EOS. Por tanto, unas condiciones otoñales más cálidas y húmedas podrían provocar un retraso en el proceso de senescencia de las hojas.
Comportamiento de SOS y EOS entre zonas ecológicas durante 2000-2010 y 2011-2019
El análisis factorial reveló que el día del año (doy) medio para el inicio de la temporada de crecimiento (SOS) de la especie fue de 160.55 (P<0.05). Los efectos significativos en el diseño factorial dependieron de la zona (D, CV, S) tanto para el SOS como para el final de la temporada de crecimiento (EOS). El factor que causó el mayor cambio en SOS y EOS (es decir, la variabilidad en doy) fue la zona ecológica en la que crece la vegetación. La interacción entre las zonas ecológicas y los períodos de tiempo analizados influyó significativamente tanto en el SOS como en el EOS (P<0.005).
Conclusiones e implicaciones
Esta investigación ha proporcionado una comprensión detallada sobre la dinámica estacional del pasto banderita [Bouteloua curtipendula (Michx.) Torr.] en diferentes zonas ecológicas del estado de Chihuahua, México, utilizando un enfoque geoestadístico respaldado por datos satelitales y observaciones fenológicas in situ. Los resultados muestran que la variabilidad climática, particularmente en temperatura y precipitación, afecta el inicio (SOS) y final (EOS) de su temporada de crecimiento. Se observó que en la zona desértica (D), el SOS ocurre antes debido a temperaturas más altas. Además, la disminución de precipitación retrasó el SOS en todas las zonas, mientras que, en otoño, las temperaturas más cálidas extendieron el EOS en la zona D. La comprensión de la dinámica estacional de esta especie es esencial para desarrollar prácticas de manejo sostenible que aseguren la resiliencia de estos ecosistemas frente al cambio climático. Los resultados obtenidos proporcionan una base científica robusta para la toma de decisiones informadas en la gestión de los recursos naturales en la región, promoviendo la sustentabilidad y la adaptación a las nuevas condiciones ambientales.
Conflicto de intereses
Los autores declaran no tener conflicto de intereses.
Literatura citada: