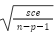 . Donde: n = número total de datos; sce= suma de cuadrados del error; sct = suma de cuadrados total; k= número de parámetros en el modelo; ln = logaritmo natural. Para AIC y BIC, el modelo con el menor valor se consideró como de mejor ajuste.
. Donde: n = número total de datos; sce= suma de cuadrados del error; sct = suma de cuadrados total; k= número de parámetros en el modelo; ln = logaritmo natural. Para AIC y BIC, el modelo con el menor valor se consideró como de mejor ajuste. https://doi.org/10.22319/rmcp.v14i2.6286
Nota de investigación
Curvas de crecimiento en bovinos Limousin de raza pura y cruzados
Joel Domínguez-Viveros a
Antonio Reyes-Cerón b
Carlos Enrique Aguirre-Calderón c*
Ricardo Martínez-Rocha d
Carlos Luna-Palomera e
Nelson Aguilar-Palma a
a Universidad Autónoma de Chihuahua. Facultad de Zootecnia y Ecología. Chihuahua, México.
b Asociación Mexicana de Criadores de Ganado Limousin. Zacatecas, México.
c Instituto Tecnológico de México. Instituto Tecnológico de El Salto. El Salto, Durango, México.
d Universidad Nacional Autónoma de México. Facultad de Estudios Superiores Cuautitlán. Ciudad de México, México.
e Universidad Juárez Autónoma de Tabasco. División Académica de Ciencias Agropecuarias. Tabasco, México.
*Autor de correspondencia: carlos.ac@salto.tecnm.mx
Resumen:
El objetivo fue el ajuste de un modelo no lineal (MNL) para evaluar la curva de crecimiento en bovinos Limousin, en pureza de raza (PRZ) y cinco grados de cruzamiento (GPZ; 1/2, 3/4, 7/8, 15/16, 31/32 de Limousin). Se analizó el peso vivo, el intervalo de peso al nacer a 500 días de edad. Se evaluaron cuatro MNL: Brody, Bertalanffy, Gompertz y logístico. Se estimaron parámetros de crecimiento: peso adulto (PAD); tasa de crecimiento (TAC); edad (EPI; meses) y peso (PPI; kg) al punto de inflexión; edad (meses; E50M) para alcanzar 50% de madurez y madurez a 15 meses (GM15). Con el MNL seleccionado en PRZ se caracterizó la curva de crecimiento en GPZ. El modelo de mejor ajuste fue Bertalanffy. El PAD para machos PRZ fue 566.1, para GPZ estuvo en el intervalo de 446.9 a 527.4; para hembras, en GPZ estuvo en el intervalo de 374.5 a 419.9, en PRZ fue 443.0. Los MNL presentaron correlaciones por debajo de -0.75 entre PAD y TAC. En vaquillas de PRZ, EPI se estimó a 3.7 con 131.2 para PPI; en GPZ, EPI y PPI estuvieron en los intervalos de 2.9 a 3.7 y 110.9 a 124.4, respectivamente. E50M para hembras, en PRZ fue a 10.6 y para GPZ en el intervalo de 8.9 a 10.5. GM15 para hembras, en GPZ el promedio fue 90.5 % en PRZ fue de 87.9 %. Los machos en PRZ alcanzan E50M a partir de 13 meses.
Palabras clave: Modelos no lineales, Parámetros de crecimiento, Bos Taurus, Cruzamiento, Heterosis.
Recibido: 10/07/2022
Aceptado: 07/11/2022
La raza Limousin, originaria de Francia, como raza pura o en esquemas de cruzamiento(1,2) presenta cualidades productivas, reproductivas y de adaptación que han permitido su distribución en gran cantidad de países y de sistemas de producción(3,4,5); además, se ha utilizado en el desarrollo de razas sintéticas(6). A México llegó por importaciones de Canadá y Estados Unidos que datan de la década de 1970; la Asociación Mexicana de Criadores de Ganado Limousin (AMCGL) se constituyó en 1989(7,8). Actualmente se distribuye en 17 estados como raza pura especialmente; aunque también es utilizada en esquemas de cruzamiento abierto y como base para la conformación de razas sintéticas, tales como Limousan (5/8 Limousin y 3/8 Angus)(9) y Brahmousin (5/8 Limousin y 3/8 Brahman)(10).
La AMCGL coordina el registro genealógico de pureza de raza y grados de pureza, así como los registros productivos que definen los criterios y objetivos de selección de la raza(7). Los datos productivos asociados al crecimiento incluyen el peso vivo al nacer, a 120, 210 y 365 días de edad, con mediciones en el intervalo de más o menos 45 días a la edad especificada. Las mediciones de peso vivo generan una distribución de observaciones a través de la vida del animal, que en su conjunto se pueden utilizar para caracterizar y evaluar la curva de crecimiento. Los modelos no lineales (MNL) caracterizan y analizan la curva de crecimiento animal con base en la interpretación biológica y aplicaciones de los coeficientes de regresión, así como parámetros de crecimiento derivados de los coeficientes de regresión(11,12,13). Los coeficientes de regresión y parámetros de crecimiento son de trascendencia en la toma de decisiones para los programas de manejo, nutrición, reproducción y mejora genética(14,15,16,17). Con base en lo anterior, el objetivo del presente estudio fue la selección y ajuste de un MNL para describir y evaluar la curva de crecimiento en bovinos Limousin de México.
La base de datos estuvo conformada por mediciones de peso vivo, en el intervalo de peso al nacer a 500 días de edad en bovinos Limousin (PRZ; pureza de raza). Para definir la curva de crecimiento se evaluaron cuatro MNL: Brody (BRO), von Bertalanffy (BER), Gompertz (GOM), y logístico (LOG); todos conformados por tres coeficientes (β1, β2 y β3) de regresión(12,13,18). En las ecuaciones de los MNL (Cuadro 1), yi representa el peso vivo (kg) medido al tiempo t; β1, es el valor asintótico cuando t tiende a infinito, interpretado como el parámetro de peso adulto (PAD); β2, es un parámetro de ajuste cuando y ≠ 0 y t ≠ 0; y β3, es la tasa de crecimiento (TAC), expresando la ganancia de peso como proporción del peso total. Los modelos BER, GOM y LOG se caracterizan por describir el crecimiento con base en una curva sigmoide, para los cuales se calculó la edad (EPI; meses) y el peso (PPI; kg) al punto de inflexión. El modelo BRO presenta una curva de crecimiento con TAC constante sin punto de inflexión. Con los coeficientes de regresión se estimaron la edad para alcanzar el 50 % de madurez (E50M), la madurez obtenida a los 15 meses (GM15) de edad(19,20), así como la correlación (rac) entre TAC y PAD.
Cuadro 1: Modelos no lineales evaluados en bovinos Limousin de raza pura y cruzados
Modelo | Ecuación | |
Logístico | yi = β1 / (1 + β2*(exp(-β3*t))) + ei | |
Bertalanffy | yi = β1*((1 - β2*(exp(-β3*t)))**3) + ei | |
Gompertz | yi = β1*(exp(-β2*(exp(-β3*t)))) + ei | |
Brody | yi = β1*(1 - β2*(exp(-β3*t))) + ei |
yi= peso vivo en kg, medido al tiempo t; β1= valor asintótico; β2= constante de integración; β3= pendiente de la curva o tasa de crecimiento.
Los análisis se realizaron para cada sexo, con el método de Gauss-Newton del procedimiento NLIN del programa para análisis estadístico SAS(21). La selección del modelo con mejor ajuste se realizó en función de(18,19): criterio de información Akaike [AIC= n*ln(sce/n) + 2k]; criterio de información Bayesiano [BIC= n*ln(sce/n) + k*ln(n)]; coeficiente de determinación [R2= (1 – (sce/sct))]; y, error estándar general o del modelo (EEG= . Donde: n = número total de datos; sce= suma de cuadrados del error; sct = suma de cuadrados total; k= número de parámetros en el modelo; ln = logaritmo natural. Para AIC y BIC, el modelo con el menor valor se consideró como de mejor ajuste.
La AMCGL administró un registro genealógico con diferentes grados de pureza (GPZ) con el objetivo de incrementar la población de bovinos Limousin a través de cruzamiento absorbente, con base en vacas cruzadas y sementales de PRZ. Con el modelo seleccionado como de mejor ajuste en la población de PRZ se caracterizó la curva de crecimiento en poblaciones definidas por cinco GPZ o generaciones: primera (PG) con ½ de Limousin; segunda (SG) con ¾ de Limousin; tercera (TG) con 7/8 de Limousin; cuarta (CG) con 15/16 de Limousin; y, quinta (QG) con 31/32 de Limousin. En el Cuadro 2 se describe la base de datos analizada en PRZ y GPZ.
Cuadro 2: Base de datos de peso vivo, analizada a través de grupos genéticos y de sexo, con mediciones en el intervalo del nacimiento hasta 500 días de edad
Sexo / Grupo | PG | SG | TG | CG | QG | Pureza |
Machos | 1963 | 1489 | 1607 | 3428 | 6224 | 31784 |
Hembras | 2220 | 2296 | 2449 | 4784 | 7382 | 35695 |
Grupos genéticos: PG, 1/2 Limousin; SG, 3/4 Limousin; TG, 7/8 Limousin; CG, 15/16 Limousin; QG, 31/32 Limousin. Pureza de raza (> 63/64 Limousin).
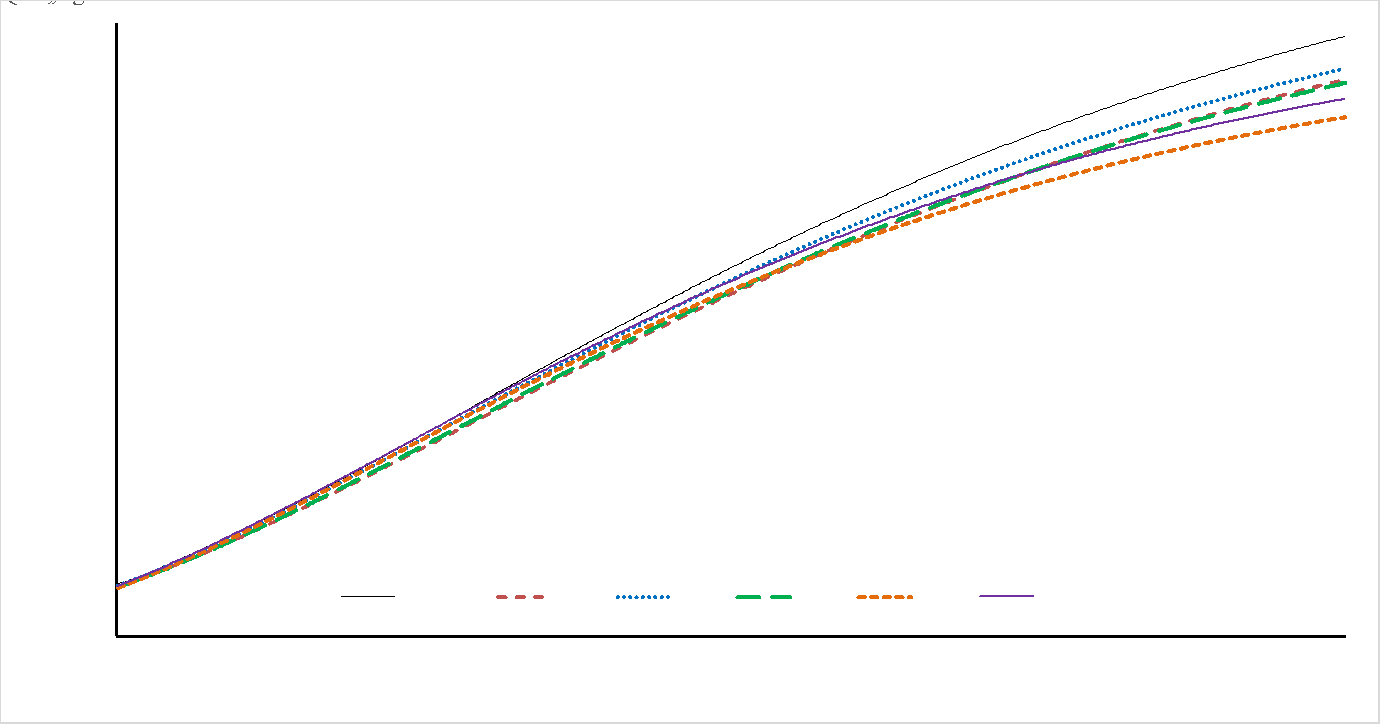
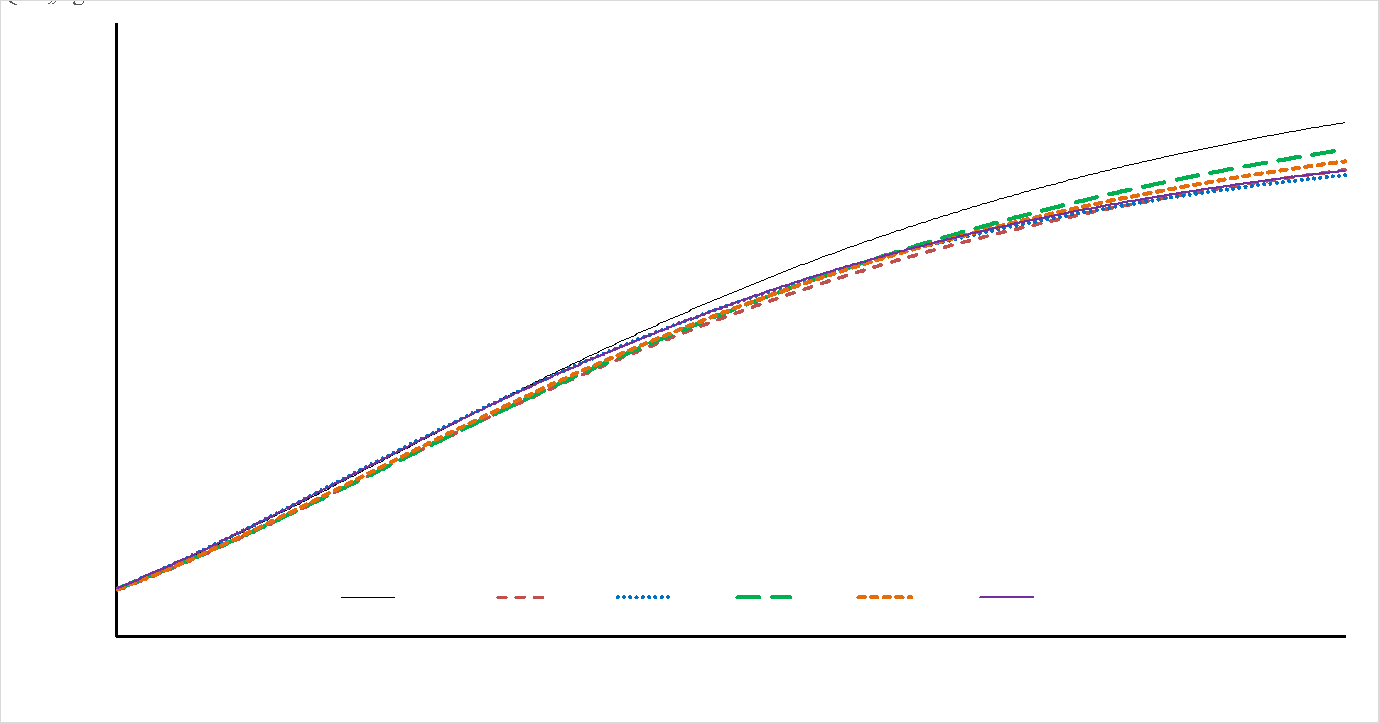
En la selección de modelos, dentro de sexo con AIC y a través de sexo con BIC, el modelo de mejor ajuste fue BER seguido de BRO y GOM; en todos los modelos el R2 fue superior al 95 % (Cuadro 3). En el Cuadro 4 se presentan los resultados para los coeficientes de regresión y parámetros de crecimiento producto de los MNL evaluados. La estimación del PAD fue mayor para PRZ vs GPZ, en contraste, la TAC fue superior en los resultados de GPZ. El esquema de mejora genética del ganado Limousin en México incluye el peso al destete ajustado a 205 días(7), con posible trascendencia en las curvas de crecimiento, dado que el punto de inflexión está ubicado en el periodo predestete. El modelo de BRO fue el segundo en la clasificación de modelos; sin embargo, presentó resultados atípicos para PAD, E50M y GM15. Todos los modelos presentaron rac debajo de -0.75 (Cuadro 4), lo cual señala que altos PAD no derivan de altas TAC. En la Figura 1 para machos y Figura 2 para hembras, se describe el crecimiento con base en el modelo BER para todos los genotipos evaluados.
Cuadro 3: Estadísticos utilizados para la selección del modelo no lineal de mejor ajuste
Estadísticos | Brody | Gompertz | Logístico | Bertalanffy |
Machos | ||||
R2 | 96.7 | 96.7 | 96.6 | 96.7 |
EEG | 40.3 | 40.3 | 40.9 | 40.3 |
AIC | 236935.8 | 237022.3 | 237856.6 | 236896.4 |
BIC | 236960.9 | 237047.5 | 237881.7 | 236921.2 |
Hembras | ||||
R2 | 96.8 | 96.8 | 96.7 | 96.8 |
EEG | 35.6 | 35.7 | 36.1 | 35.6 |
AIC | 255208.7 | 255253.0 | 256169.4 | 255127.7 |
BIC | 255234.2 | 255278.4 | 256194.9 | 255153.1 |
AIC= criterio de información Akaike; BIC= criterio de información Bayesiano; R2= coeficiente de determinación; EEG= error estándar general o del modelo.
Cuadro 4: Coeficientes de regresión y parámetros de crecimiento derivados de los modelos no lineales evaluados en bovinos Limousin de raza pura y cruzados
ítem | β1 | β2 | β3 | rac | EPI | PPI | E50M | GM15 |
Machos de raza pura con todos los modelos no lineales evaluados | ||||||||
Brody | 1645.9 | 0.9778 | 0.000618 | -0.99 | -- | -- | 36.2 | 26.0 |
Gompertz | 491.0 | 2.5475 | 0.00583 | -0.92 | 5.3 | 180.6 | 7.4 | 83.1 |
Logístico | 408.2 | 8.8538 | 0.0117 | -0.76 | 6.2 | 204.1 | 6.2 | 98.5 |
Bertalanffy | 566.1 | 0.5949 | 0.00400 | -0.96 | 4.8 | 167.7 | 13.0 | 81.2 |
Hembras de raza pura con todos los modelos no lineales evaluados | ||||||||
Brody | 715.1 | 0.9508 | 0.00151 | -0.99 | -- | -- | 14.2 | 51.8 |
Gompertz | 402.9 | 2.396 | 0.00656 | -0.90 | 4.4 | 148.2 | 6.3 | 88.2 |
Logístico | 352.3 | 8.0113 | 0.0124 | -0.71 | 5.6 | 176.1 | 5.6 | 99.1 |
Bertalanffy | 443.0 | 0.5666 | 0.00477 | -0.95 | 3.7 | 131.2 | 10.6 | 87.9 |
Machos en grado de pureza con el modelo Bertalanffy | ||||||||
PG | 527.4 | 0.5896 | 0.00394 | -0.97 | 4.8 | 156.3 | 13.1 | 80.7 |
SG | 522.4 | 0.5858 | 0.00418 | -0.96 | 4.5 | 154.8 | 12.3 | 83.0 |
TG | 514.5 | 0.5876 | 0.00410 | -0.96 | 4.6 | 152.4 | 12.6 | 82.3 |
CG | 446.9 | 0.5705 | 0.00481 | -0.95 | 3.7 | 132.4 | 10.5 | 88.1 |
QG | 467.1 | 0.5724 | 0.00469 | -0.96 | 3.8 | 138.4 | 10.8 | 87.3 |
Hembras en grados de pureza con el modelo Bertalanffy | ||||||||
PG | 391.4 | 0.5563 | 0.00511 | -0.94 | 3.3 | 115.9 | 9.7 | 90.0 |
SG | 374.5 | 0.5446 | 0.00563 | -0.94 | 2.9 | 110.9 | 8.7 | 92.6 |
TG | 419.9 | 0.5619 | 0.00476 | -0.95 | 3.7 | 124.4 | 10.5 | 87.9 |
CG | 399.2 | 0.5572 | 0.00509 | -0.95 | 3.4 | 118.3 | 9.8 | 89.9 |
QG | 379.9 | 0.5471 | 0.00551 | -0.94 | 3.0 | 112.5 | 8.9 | 92.1 |
Grados de pureza: PG, 1/2 Limousin; SG, 3/4 Limousin; TG, 7/8 Limousin; CG, 15/16 Limousin; QG, 31/32 Limousin. Coeficientes de regresión: β1, β2 y β3. Donde: β1, es el valor asintótico, interpretado como el parámetro de peso adulto; β2, es un parámetro de ajuste; β3, es la tasa de crecimiento, expresando la ganancia de peso como proporción del peso total. Edad (EPI; meses) y el peso (PPI; kg) al punto de inflexión. E50M, edad para alcanzar el 50 % de madurez. GM15, grado de madurez (%) a 15 meses de edad. rac, correlación entre β1 y β3.
Figura 1: Curvas de crecimiento para machos Limousin. Pureza, animales de raza pura; PG, 1/2 de Limousin; SG, 3/4 de Limousin; TG, 7/8 de Limousin; CG, 15/16 de Limousin; QG, 31/32 de Limousin
Figura 2: Curvas de crecimiento para hembras Limousin. Pureza, animales de raza pura; PG, 1/2 de Limousin; SG, 3/4 de Limousin; TG, 7/8 de Limousin; CG, 15/16 de Limousin; QG, 31/32 de Limousin
En bovinos Limousin de raza pura, Igarzabal et al(3) en tres sistemas de producción reportaron como modelo de mejor ajuste GOM. En esquemas de cruzamiento de Limousin con Angus, Hereford y MARC III, Zimmermann et al(17) utilizaron el modelo de BRO para caracterizar la curva de crecimiento y evaluar el peso vivo a la madurez. En bovinos Limousin x Friesian, representaron el crecimiento con base en el modelo GOM(22). En la raza Madrasin, producto del cruzamiento de Limousin con Madura, el crecimiento presentó una curva tipo sigmoide, caracterizada con el modelo LOG(23). Curvas de crecimiento evaluadas con el modelo BER, fueron reportadas en bovinos Holstein(24), Pirenaica y Blonda(3).
En México, diversos estudios han discutido contrastes en el tipo de curva de crecimiento a través de razas. Para curvas de crecimiento sin punto de inflexión, Domínguez-Viveros et al(25) en cinco razas de cebú en ganadería tropical, reportaron que los MNL de mejor ajuste fueron Brody, Meloum III y Mitscherlich; caso particular del modelo BRO, fue seleccionado con mejor ajuste en vacas Romosinuano(20), en bovinos Tropicarne(19) y Salers(26). Para curvas de crecimiento tipo sigmoide, Contreras et al.(27) en vacas Jersey, Holstein y cruzas Jersey con Holstein los MNL seleccionados fueron GOM, LOG y BER, respectivamente; el modelo BER se ha reportado en bovinos Hereford(26).
La incorporación de vaquillas de reemplazo a la fase reproductiva es de trascendencia para el progreso genético y rentabilidad del hato. Este procedimiento transcurre en tres etapas(28): inicia maduración de la hipófisis detonada a cierta edad y peso; seguida con el desarrollo de los ovarios y crecimiento corporal; maduración del útero como consecuencia del desarrollo pituitario y su influencia hormonal sobre el crecimiento corporal y actividad ovárica, permite a la vaquilla el empadre y desarrollar la gestación. En diversos estudios se ha analizado la influencia de los parámetros de crecimiento sobre variables reproductivas(20,24,29); de esta forma, el punto de inflexión se ha asociado(13,30,31) con el inicio de la fase reproductiva. La edad al primer parto es un indicador del tiempo que tarda un animal en alcanzar su madurez sexual y reproducirse por primera vez, el empadre alrededor de los 15 meses, con edad al primer parto en torno a los 24 meses, tiene efectos positivos en la longevidad y productividad de la vaca(32,33). Con base en el modelo BER, se observan diferencias en las hembras de GPS vs PRZ para los componentes de la curva de crecimiento (Cuadro 4), las cuales se pueden atribuir a las diferencias genéticas a través de razas y los efectos de heterosis, producto del esquema de cruzamiento. En vaquillas de PRZ, la EPI se estimó a 3.7 meses con 131.2 kg para PPI; en GPZ, la EPI y el PPI estuvieron en los intervalos de 2.9 a 3.7 meses y 110.9 a 124.4 kg, con valores promedio de 3.3 y 116.4, respectivamente. Para E50M en hembras, en PRZ se estimó a 10.6 meses y para GPZ estuvo en el intervalo de 8.9 a 10.5 meses, con un valor promedio de 9.5. Para GM15 en hembras, en GPZ el valor promedio fue de 90.5 % y la estimación en PRZ fue de 87.9 %. En contraste, para hembras de otras poblaciones y con base en el modelo BER: Contreras et al(27) para Holstein, Jersey y cruzas estimaron EPI (meses) y PPI (kg) en los intervalos de 7.4 a 9.8 y 115.0 a 151.7, respectivamente; Domínguez-Viveros et al(25) en cinco razas de cebú, reportaron estimaciones de EPI y PPI en los intervalos de 3.9 a 11.7 y 107.2 a 230.9, respectivamente; en Romosinuano(20), Tropicarne(19) y Siboney(29) los resultados de EPI – PPI fueron de 15.5 – 132.5, 7.7 – 180.5 y 5.9 – 152.4, respectivamente.
Con respecto a los machos, la selección de sementales se realiza en PRZ y se incorporan a la reproducción a partir del año de edad; no obstante, reducir la edad de incorporación a la reproducción reduce el intervalo generacional y trasciende en el progreso genético(34). La curva de crecimiento puede incidir en el desarrollo de la fase reproductiva; en razas Bos taurus, los eventos fisiológicos asociados a la reproducción inician a los seis u ocho meses; la madurez y capacidad reproductiva se precisan por la calidad del semen, con variaciones por efectos del peso vivo, tasa de crecimiento, circunferencias escrotal, entre otros factores(35). Los resultados señalan que los machos en PRZ alcanzan el 50 % de madurez a partir de los 13 meses, con valores superiores al 80 % a partir de los 15 meses (Cuadro 4). En contraste(26), en bovinos Hereford y Salers se reportaron grados de madurez de 68.2 % y 76.6 %, al año, respectivamente. Por otro lado, los indicadores de la curva de crecimiento están asociados a la rentabilidad en la producción; la TAC tiene efecto en la edad y peso al sacrificio; grado de madurez trascendencia en la eficiencia y composición de la canal(16,22). Diferencias y derivaciones de la curva de crecimiento con relación a la producción, para machos de raza pura y diversos cruzamientos, han sido valoradas por diversos autores(15,36,37).
El modelo de mejor ajuste fue von Bertalanffy, el cual describió una curva de crecimiento tipo sigmoide, con diferencias en los parámetros de crecimiento a través de los genotipos evaluados. Las estimaciones del punto de inflexión están en el contexto del crecimiento predestete.
Agradecimientos
Se agradece a la Asociación Mexicana de Criadores de Ganado Limousin por facilitar la base de datos analizada.
Literatura citada: