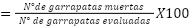
https://doi.org/10.22319/rmcp.v14i3.6265
Artículo
Evaluación in vitro del potencial acaricida de Beauveria bassiana DS3.17 sobre la garrapata común (Rhipicephalus microplus) en Oaxaca, México
Jared Martínez-García a
José Abad-Zavaleta b
María de Jesús García-Gómez b
Oscar Núñez-Gaona b*
a Universidad del Papaloapan. División de Estudios de Posgrado, Maestría en biotecnología, Oaxaca, México.
b Universidad del Papaloapan. Instituto de Biotecnología, Centro de Investigaciones Científicas. Oaxaca, México.
*Autor de correspondencia: oscarnzg@hotmail.com
Resumen:
La garrapata Rhipicephalus microplus causa daños a la ganadería bovina, tiene un gran impacto económico e influye significativamente sobre la productividad y competitividad comercial del sector, principalmente en zonas tropicales y subtropicales del país, generando pérdidas anuales millonarias. Actualmente, el control del ectoparásito empleando agentes biológicos (hongos entomopatógenos, principalmente) tiene un elevado potencial para reducir las poblaciones de las garrapatas. El objetivo del presente estudio fue evaluar in vitro el potencial acaricida de Beauveria bassiana DS3.17 sobre hembras ingurgitadas de R. microplus, mediante inmersión del ácaro en diferentes concentraciones de conidios del hongo para determinar los efectos sobre los parámetros reproductivos (peso total de huevos, porcentaje de eclosión, índice de ovoposición y nutrición). Las concentraciones evaluadas de B. bassiana DS3.17 evidenciaron un efecto acaricida de hasta un 93.2 % en adultos y un cambio en el comportamiento de los parámetros biológicos de las hembras adultas, inhibiendo la ovoposición entre 75.84 y 79.60 %. En cuanto al índice de reproducción estimada se obtuvo un control del 45.23 al 83.79 %, el índice de nutrición fue de 10.21 y 10.67 %. El estudio mostró un cambio en el comportamiento de los parámetros reproductivos cuando se utilizó B. bassiana DS3.17, y cada uno de los resultados mostraron diferencias significativas; las concentraciones de conidios afectaron la reproducción del parásito respecto al control, por lo que B. bassiana DS3.17 presentó un potencial acaricida para el control de la garrapata.
Palabras clave: Control biológico, Beauveria bassiana, Rhipicephalus microplus.
Recibido: 21/06/2022
Aceptado: 09/03/2023
Introducción
Los daños causados por Rhipicephalus microplus al ganado bovino tienen un gran impacto económico e influyen significativamente sobre la productividad y competitividad comercial del sector en las zonas tropicales y subtropicales del país, generando pérdidas anuales de 573 millones de dólares(1,2,3).
La aplicación de ixodicidas sintéticos juega un papel importante en el control de esta plaga, siendo el método más utilizado para disminuir sus poblaciones(4,5). Sin embargo, el uso indiscriminado de estos productos genera un impacto negativo al ambiente, contribuyendo al surgimiento de plagas secundarias, disminución de enemigos naturales y desarrollo de resistencia por parte de los ácaros(2,6,7).
Por otra parte, el control biológico que emplea hongos entomopatógenos; es una de las múltiples estrategias enfocadas a solucionar la problemática de los ixodicidas sintéticos, esta metodología va en aumento debido a la conciencia que ha adquirido la sociedad sobre la seguridad ambiental y la salud humana, aunado al incremento en el costo al aplicar el control químico(8). Además, los agentes de control biológico destacan por su amplia distribución, riesgo bajo para la salud animal y humana, compatibilidad con el ambiente y alta virulencia(1,4).
Cepas de los géneros Beauveria bassiana y Metarhizium anisopliae han sido evaluadas contra garrapatas; debido a su virulencia, mecanismo de infección, mortalidad sobre adultos, larvas, por alterar la eclosión de los huevos y generar cambios en el metabolismo lipídico de su hospedero(9-12). A pesar de los resultados favorables obtenidos in vitro y en campo, estos microorganismos muestran un comportamiento errático debido a que su actividad se reduce al aplicarse en campo, por lo que su adaptación a las condiciones ambientales es un factor determinante sobre su patogenicidad y virulencia. Además, se ha descrito que algunos hongos entomopatógenos son más susceptibles a factores abióticos, particularmente temperatura y radiación ultravioleta(13). En este sentido, los hongos endémicos están mejor adaptados a dichas condiciones(1,14) y juegan un papel importante en el control natural de poblaciones plaga. El objetivo del presente trabajo fue determinar el efecto acaricida de B. bassiana DS3.17 sobre la viabilidad y los parámetros reproductivos de hembras ingurgitadas de R. microplus.
Material y métodos
Microorganismo
Se empleó B. bassiana DS3.17 perteneciente a la colección de cepas del cuerpo académico Biotecnología Sustentable de la Universidad del Papaloapan Tuxtepec Oaxaca, México. La cepa fue aislada de Rancho Grande (latitud, 17.844256, longitud, -96.333192), San Juan Bautista Valle Nacional, Oaxaca, México. Todas las coordenadas se obtuvieron empleando un Navegador de mano modelo eTrex Legend (Garmin Ltd, EE. UU.). La cepa se incubó a 25 °C durante 14 días en medio agar Dextrosa Sabouraud al 4% (p/v) suplementado con 0.05% (p/v) de extracto de levadura (Bioxon, México). La producción del inóculo fúngico se realizó por fermentación bifásica empleando la metodología descrita por López-Sosa et al(15). Al final de la fermentación, los conidios se recuperaron mediante tamizado (300 µm, TEST SIEVE Alemania), se suspendieron en agua destilada estéril (1:10 g ml-1) hasta una concentración final de 4.2x109 conidios ml-1; a partir de ésta se realizaron diluciones para obtener suspensiones con concentraciones de 1x106,1x107 1x108 y 1x109 conidios ml-1 que fueron utilizadas para los bioensayos. Los valores de las diluciones se terminaron con base en la literatura; buscando cubrir el intervalo de concentraciones empleadas en ella.
Las garrapatas adultas (R. microplus) se colectaron en el rancho la Guadalupe ubicado en la comunidad de Peñarrubia (17º 50’ 39’’N, 96º 19’ 59’’O) Tuxtepec, Oaxaca, México. Los ejemplares se depositaron en viales de polipropileno con capacidad de 50 ml, se descartaron aquellos que se consideraron no viables (pérdida de extremidades, ausencia de movilidad o la muerte del organismo) para el estudio. Las garrapatas seleccionadas se desinfectaron con hipoclorito de sodio al 1% (v/v) durante 1 min, realizando tres lavados consecutivos con agua destilada estéril, posteriormente se colocaron en papel absorbente estéril para eliminar el exceso de agua. Finalmente, se incubaron en cámara húmeda a 26 ± 1 °C y humedades relativas de 70 a 80 %, monitoreándolas durante tres días para rectificar su viabilidad. Al término del periodo, las hembras ingurgitadas fueron pesadas y clasificadas en grupos de 10 individuos para realizar los bioensayos.
Bioensayos
El efecto acaricida de B. bassiana DS3.17 se evaluó con la metodología de inmersión de adultos utilizando cuatro concentraciones de conidios (1x106, 1x107, 1x108, y 1x109 conidios ml-1) y un testigo (agua destilada estéril); en cada una se sumergieron 10 hembras completamente ingurgitadas con peso aproximado entre 200 y 300 mg durante 3 min. Posteriormente, las garrapatas se colocaron en papel absorbente estéril hasta eliminar el exceso de humedad. Cada grupo se colocó en cámaras húmedas, a 26 ± 1 °C y de 70 a 80 % de humedad relativa, realizándose observaciones cada 48 h durante 23 días para evaluar la mortalidad diaria. Los bioensayos se concluyeron cuando el grupo testigo sobrepasó el 10 % de mortalidad(16).
Efecto acaricida de B. bassiana DS3.17 sobre el potencial reproductivo de R. microplus
El efecto acaricida del hongo sobre los parámetros reproductivos de la garrapata fue evaluado en cinco grupos(replicas), más 1 control de 10 garrapatas para cada concentración del hongo; mediante la metodología descrita en el apartado anterior. Después de 10 días de realizar la inmersión de los ácaros en las diluciones de conidios, se retiraron los huevos de cada uno de los grupos y se pesaron; de la cantidad colectada se tomaron alícuotas de 50 mg por cada grupo y se incubaron en cámaras húmedas hasta su eclosión bajo las condiciones ya descritas. Los parámetros evaluados fueron peso inicial y final de la garrapata, peso de los huevos, huevos eclosionados; con estos se determinó el porcentaje de eclosión (%E), índice de nutrición (IN), eficiencia reproductiva (ER) y ovoposición (OP), usando las ecuaciones propuestas por Bennett(17). A partir de los parámetros estimados se calcularon los índices de inhibición de la ovoposición (IOP) y la inhibición de la eficacia reproductiva (IER).
Ecuaciones
Porcentaje de mortalidad:
Ovoposición estimada: 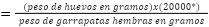
*constante del número de huevos en un gramo de masa de huevos.
Índice de nutrición: 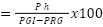
Ph= peso de los huevos; PGI= peso de las garrapatas ingurgitadas; PRG= peso residual de las garrapatas.
Inhibición de la ovoposición:
OPT= ovoposición del tratamiento; OPC= ovoposición del grupo testigo.
Eficiencia reproductiva: 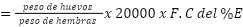
F.C.= fracción centecimal
Inhibición de la eficiencia reproductiva:  RET= Efectividad reproductiva del tratamiento; REC= efectividad reproductiva del control.
RET= Efectividad reproductiva del tratamiento; REC= efectividad reproductiva del control.
Análisis estadístico
Para el análisis de mortalidad de adultos los datos se transformaron con la ecuación: Y=asen (√(p ), donde: “Y” es la mortalidad transformada, “asen” es el arcoseno y “p” es la mortalidad en proporción(18).
El análisis estadístico se realizó utilizando el paquete estadístico, “Rcmdr”, “drc”. “MASS”, “ggplot2” y “ecotox” del software libre “R” (R Development Core Team. EE. UU.). A los datos obtenidos de la mortalidad, peso final y peso de huevos, se les realizó un análisis de varianza y comparación entre medias con la prueba de Tukey (α≤0.05). Los datos no paramétricos como el porcentaje de eclosión, índice de nutrición, inhibición de ovoposición y efectividad reproductiva se analizaron con la prueba de Kruskal-Wallis y la prueba de comparación de Student-Newman-Keuls (α≤0.05).
Resultados
La mortalidad diaria producida por B. bassiana DS3.17 sobre hembras ingurgitadas de R. microplus (Figura 1) inició al tercer día después de la inmersión para todas las concentraciones evaluadas, presentando porcentajes de mortalidad de 1.66 % (1x106 conidios ml-1), 3.33 % (1x107 conidios ml-1), 5.83 % (1x108 conidios ml-1), 6.66 % (1x109 conidios ml-1), en el grupo testigo el inicio de la mortalidad se observó al día 11. La velocidad de mortalidad fue directamente proporcional a la concentración de conidios. La máxima mortalidad se alcanzó el día 23 (97.5 y 98.3 %) con las concentraciones de 1x108 y 1x109 conidios respectivamente, al mismo tiempo las mortalidades con concentraciones de 1x106 y 1x107 conidios fueron 71.6 y 79.16 %, para el testigo; a este tiempo se alcanzó un porcentaje de mortalidad de 18.33 %. Los valores de mortalidad obtenidos con las concentraciones de 1x108 y 1x109 conidios no mostraron diferencias significativas; sin embargo, sí las mostraron con el control y el resto de las concentraciones.
Figura 1: Curva de mortalidad diaria (23 días) de hembras ingurgitadas adultas de R. microplus después de la inmersión en suspensiones de conidios de B. bassiana DS3.17 a diferentes concentraciones control. 1x106. 1x107 1x108 1x109
Efecto acaricida de B. bassiana DS3.17 sobre el potencial reproductivo de R. microplus
Las concentraciones evaluadas de B. bassiana DS3.17 tuvieron un efecto sobre los parámetros reproductivos de las hembras adultas de R. microplus (Cuadro 1). Los pesos de los huevos sumergidos en las concentraciones 1x108 y 1x109 conidios ml-1 (0.200 y 0.131 g respectivamente) fueron significativamente menores (α≤0.05) al obtenido con el testigo (0.650 g) y con la concentración de 1x107 conidios ml-1(0.588 g). La misma tendencia se observó para el índice de nutrición. En el caso del porcentaje de eclosión se observó una relación inversamente proporcional con respecto a la concentración de conidios del hongo, también no se encontraron diferencias significativas para las concentraciones de 1x108 y 1x109 conidios.
Cuadro 1: Efecto de la concentración de conidios de B. bassiana DS3.17 sobre peso inicial, peso final, peso de huevecillos, eclosión e índice de nutrición de hembras adultas de R. microplus
Concentración (conidios ml-1) | Peso inicial (g) | Peso final (g) | Peso huevecillo (g) | Índice de nutrición (%) | Eclosión (%) |
Testigo | 0.2127±0.053 | 0.333±0.045d | 0.651±0.123a | 37.08±4.93a | 87.86±4.46a |
1x107 | 0.2504±0.047 | 0.547±0.045c | 0.588±0.121a | 29.93±4.97b | 62.70±8.24b |
1x108 | 0.2790±0.054 | 0.814±0.040b | 0.201±0.08b | 10.18±3.69c | 31.53±4.74c |
1x109 | 0.2321±0.063 | 0.964±0.017a | 0.131±0.069b | 9.63±2.27c | 28.29±3.97c |
abc Letras iguales en la misma columna no presentan diferencias significativas (P>0.05).
En el Cuadro 2 se presentan; la ovoposición estimada, porcentaje de inhibición de la ovoposición, eficacia reproductiva (ER) e inhibición de la eficacia reproductiva (IER) para cada una de las concentraciones evaluadas; en donde se observa que los valores de la ovoposición presentaron una disminución inversamente proporcional a las concentraciones evaluadas, siendo el control (6,174 huevos) el grupo con la mayor cantidad de huevos estimados a diferencia de las concentraciones de 1x108 (1,434) y 1x109 conidios ml-1 (1,083). En la variable ovoposición se observó porcentajes de inhibición mayores al 40 % con las concentraciones evaluadas; siendo la concentración de 1x109 conidios la que presentó el mayor porcentaje de inhibición (94.36 %). Al comparar las medias obtenidas se encontraron diferencias significativas entre las concentraciones mayores (1x108 y 1x109 conidios) y la menor (1x107 conidios), de igual manera los valores de las concentraciones evaluadas fueron significativamente diferentes a los valores obtenidos en el grupo testigo.
Cuadro 2: Efecto de la concentración de conidios de B. bassiana DS3.17 sobre los parámetros de ovoposición y potencial reproductivo
Concentración (conidios ml-1) | Ovoposición estimada | Inhibición de ovoposición | Eficacia reproductiva | Inhibición de la eficacia reproductiva | |
Testigo | 6174±889c | 0c | 5427±628c | 0c | |
1x107 | 4689±530b | 45.99±7.33b | 2944±602b | 45.99±7.33b | |
1x108 | 1434±432a | 91.60±3.19a | 460.9±204.4a | 91.60±3.19a | |
1x109 | 1083±667a | 94.36±2.15a | 310.8±134.5a | 94.36±2.15a | |
abc Letras iguales en la misma columna no presentan diferencias (P>0.05).
Los valores calculados para el índice de eficacia reproductiva fueron de 32,640 larvas viables (1x109 conidios ml-1), lo que significó un porcentaje de control del 83.79 %. Para la concentración 1x108 conidios ml-1 se cuantificaron 44,074 larvas, lo que representó un 81.59 % de control. Finalmente, para la menor concentración de 1x107 conidios, los valores fueron 299,546 larvas viables y 45.23 % (Cuadro 2). Al realizar la comparación de medias de los valores obtenidos para la inhibición de la eficacia reproductiva se encontraron diferencias significativas con respecto al testigo y entre las concentraciones evaluadas.
Discusión
Los valores de mortalidad obtenidos con las concentraciones de 1x109 y 1x108 conidios ml-1 fueron similares a los reportados por García-Corredor et al(19), quienes con una concentración de 1x108 conidios obtuvieron un porcentaje de mortalidad del 90 % a los 21 días con B. bassiana, observando el inicio de la mortalidad después del día 4. De igual manera, Alcalá-Gómez et al(20), obtuvieron del 7 al 12 % de mortalidad con aislados de B. bassiana a los 5 días postratamiento, además la mortalidad máxima varió entre 84 y 100 % al día 20, periodo que coincide con lo presentado en este trabajo, siendo la cepa de B. bassiana 115 la que produjo el 100 % de mortalidad en colonias de garrapatas resistentes a ixodicidas.
Por otra parte, Tofiño-Rivera et al(4) reportaron el 100 % de mortalidad en el día 7 con una concentración de 1x108 conidios , mientras que con 1x106 conidios obtuvieron 90 % de mortalidad al día 10. Broglio-Forti et al(21) observaron el 100 % de mortalidad de garrapatas con concentraciones de 1x108 y 1x109 conidios en un periodo de 14 días. De igual manera, Del Pozo et al(22) reportaron valores de mortalidad de 87.2 a 96.4 % después de 12 días. En este sentido, en el presente trabajo se alcanzaron porcentajes de mortalidad similares, pero con tiempos más prolongados. Lo anterior pudo deberse a la complejidad distintiva de los procesos fisiológicos y enzimáticos de los hongos entomopatógenos, que están influenciados por factores como la variación en la excreción enzimática durante el proceso de infección, así como la variabilidad genética entre las cepas del mismo género, que favorece la expresión diferencial de genes de virulencia o específicos(23,24,25).
Con relación al efecto sobre los parámetros reproductivos, los valores obtenidos fueron similares a los presentados por Pulido-Medellín et al(26) quienes determinaron el efecto de M. anisopliae sobre R. microplus reportando que el índice reproductivo se redujo hasta un 91 %. Por otra parte, Alcalá-Gómez et al(20) enfatizaron que cepas de Metarhizium como la Ma136 inhibieron el 73 % (colonia endémica) y 64 % (garrapata “Media joya”) de la ovoposición, interrumpiendo la eclosión en 73 y 86 %, respectivamente, con respecto al testigo. Con base en lo anterior se puede afirmar que el comportamiento de la cepa evaluada fue similar al de otros hongos entomopatógenos; esto se debe a la secreción de proteínas que presentan relaciones ortólogas entre los hongos entomopatógenos, se ha considerado que la actividad entomopatógena está relacionada con proteínas que han evolucionado a partir de un ancestro en común(27,28). En estudios de B. bassiana, Fernández-Ruvalcaba et al(9) mencionan porcentajes de 36 a 83 % de inhibición de la ovoposición con una reducción estimada en la reproducción de 46 a 95 % con diferentes cepas, siendo B. bassiana Bb-5J5 la que presentó un mayor porcentaje de control para los parámetros descritos, con concentraciones de 5x107 conidios ml-1. Ren et al(29) encontraron que el índice de eficacia reproductiva se vio afectado por cepas de Beauveria (BbAT01, BbAT03 y BbAT13) y Metarhizium (MaAT04), observando que a mayor concentración de conidios se incrementó la reducción en la eficacia reproductiva de R. microplus, tendencia también observada en este trabajo. La reducción en la ovoposición podría deberse a la infección del ovario de la garrapata como lo indicaron Paulo et al(30); sin embargo, el desequilibrio metabólico generado durante el proceso de infección por el entomopatógeno juega un papel importante, debido a la producción de derivados de especies reactivas de oxígeno que son capaces de causar daños a diversos componentes celulares(24,31).
Así mismo, al comparar los resultados de este trabajo con el efecto de B. bassiana Bb115 sobre los parámetros reproductivos de dos colonias de garrapatas con diferente susceptibilidad a ixodicidas(20) se reportaron porcentajes de inhibición similares a los obtenidos para B. bassiana DS3.17, el índice de ovoposición fue de 0.23 para la colonia nativa y 1.94 para la colonia “media joya” (resistente a ixodicidas) valores que representaron una reducción de la ovoposición del 98 y 79 % respectivamente. Por otro lado, el índice de reproducción para las otras colonias fue de 0.11 y 0.49 para cada una, valores que significaron porcentajes de control del potencial reproductivo del 98 y 89 %.
Bernardo et al(5) obtuvieron índices de nutrición con Beauveria bassiana del 58.89 al 59.32 % para las cepas IP361 y CG307, estos valores representaron entre 35,086 y 182,061 larvas viables significando porcentajes de control de 63.29 a 92.92 %.
Por otra parte, la virulencia de los hongos con potencial para regular poblaciones de garrapatas depende de su susceptibilidad a factores como temperatura, tiempo de exposición a radiación UV(1,5,8), el origen de la cepa(32,33) y en algunos casos concentraciones elevadas de conidios aumentaron la mortalidad incrementando el control del insecto plaga(34). Esto último fue observado con la cepa DS3.17 en este trabajo, donde la concentración mayor (1x109 conidios ml-1) fue más eficaz en el control de R. microplus.
Conclusiones e implicaciones
B. bassiana DS3.17 tuvo un efecto significativo de control sobre los parámetros reproductivos de R. microplus; observándose que los índices de ovoposición y eclosión fueron inversamente proporcionales a la concentración de conidios. Esto incrementa el interés para el uso de esta cepa en el control biológico de R. microplus en ganado de la región, disminuyendo así el uso de ixodicidas. Los resultados obtenidos, abren la posibilidad de modelar el efecto de B. bassiana D3.17 sobre los parámetros reproductivos de la garrapata, optimizando los más relevantes para su control.
Agradecimientos y conflictos de interés
Jared Martínez García agradece al Consejo Nacional de Ciencia y Tecnología (CONACYT) por la beca de Maestría otorgada (942190). Los autores agradecen al Ing. Genaro Arana por la atención y disponibilidad durante la colecta de los ectoparásitos en sus instalaciones.
Todos los autores manifiestan no tener conflictos de interés en la realización del presente trabajo.
Literatura citada:





