https://doi.org/10.22319/rmcp.v13i3.6103
Revisión bibliográfica
Comportamiento de ingestión y consumo de forraje por vacas en pastoreo en clima templado. Revisión
Juan Daniel Jiménez Rosales a
Ricardo Daniel Améndola Massiotti a*
a Universidad Autónoma Chapingo. Departamento de Zootecnia, Posgrado en Producción Animal. km. 38.5 Carretera México-Texcoco, 56230, Chapingo, Estado de México. México.
* Autor de correspondencia: r_amendola@yahoo.com
Resumen:
El objetivo fue revisar, con base en publicaciones predominantemente recientes, el conocimiento sobre los componentes del comportamiento de ingestión (CI) de vacas que pastorean en clima templado, y su relación con las características de las praderas que regulan el consumo diario de forraje (CF). Los componentes del CI que median el CF son masa de bocado (MB, g MS bocado-1), tasa de bocados (TB, bocados min-1), tasa de consumo (TC, g MS min-1) y tiempo de pastoreo (TP, min día-1). La masa, altura y densidad del forraje de las praderas afectan la MB y consecuentemente, el CF. La altura de la pradera se relaciona con los componentes del CI y es útil para evaluar el CF. Con base en estudios en praderas templadas en estado vegetativo, se destaca que el CF de vacas aumenta con incrementos en altura de la pradera, porque cosechan bocados de mayor MB, lo que les permite obtener altas TC. Pero, hay evidencia de que la TC puede disminuir en praderas demasiado altas; para procesar bocados más grandes, las vacas reducen su TB y ejecutan mayor cantidad de movimientos mandibulares compuestos y de masticación. Por el contrario, en praderas cortas las vacas aumentan su TB y TP, para remediar la reducción en la TC debida a la cosecha de bocados de menor peso, aunque esto no compensa completamente la disminución de TC. Por lo anterior, para mantener altas TC las vacas no deben ser forzadas a consumir forraje a altas intensidades de pastoreo.
Palabras clave: Tasa de consumo, Tiempo de pastoreo, Tasa de bocados, Masa de bocado, Altura de pradera.
Recibido: 28/11/2021
Aceptado: 22/02/2022
Introducción
El consumo de materia seca (MS) y la digestibilidad de la dieta determinan el suministro de nutrientes a vacas estabuladas o en pastoreo, y en consecuencia afectan su producción. Por ello, la comprensión de los factores que afectan el consumo en el corto plazo de forraje por vacas en pastoreo es importante para la gestión del pastoreo, más aún cuando el consumo y, por ende, la producción de vacas en pastoreo es menor que en vacas estabuladas.
La ingesta de forraje por bovinos en pastoreo se puede evaluar a partir del comportamiento de ingestión que exhiben durante el forrajeo, ya que adaptan ese comportamiento en función de características de la pradera(1) y diversos factores como la composición química del forraje (fibra por detergente neutro, carbohidratos solubles y proteína cruda), los estímulos de los productos de la fermentación ruminal (N-NH3 y ácidos grasos volátiles), las hormonas del hambre (grelina), la saciedad (leptina y melatonina), el llenado del rumen(2), las consecuencias post-ingestivas ocasionadas por el contenido de metabolitos secundarios en las plantas(3), la suplementación y el estado fisiológico y nutricional(4). Sin embargo, en pastoreo la masa de bocado junto con características de la pradera como; masa, altura y desaparición del forraje son factores que median el consumo y en un estudio explicaron 78 % de las variaciones en la ganancia de peso de bovinos(5). Corroborando este resultado, en un metaanálisis de publicación reciente(6), que involucró 103 publicaciones con 278 experimentos, se confirmó que la masa de bocado (MB) es un componente fundamental del comportamiento de ingestión (CI) en pastoreo; ya que es sensible a las principales características del dosel de la pradera y es factor determinante para la tasa de consumo (TC) y el consumo de forraje (CF).
Por la importancia que tiene el consumo de forraje en la producción animal en pastoreo y debido a los efectos de las características del pastizal inducido (pradera) en el comportamiento de ingestión, el propósito de esta revisión es caracterizar los componentes del comportamiento de ingestión de vacas en pastoreo en clima templado, con base en resultados de investigaciones principalmente recientes, realizadas en su mayoría en praderas de clima templado homogéneas (monofitas, con escasa variación espacial en los ejes vertical y horizontal). Estos estudios, han permitido avanzar en la comprensión de las relaciones funcionales entre características de las praderas con las dimensiones del bocado y la TC(7). La predominancia del estudio del comportamiento ingestivo de bovinos en praderas homogéneas se debe a la mayor dificultad en la metodología para evaluar dicho comportamiento en vegetaciones más heterogéneas, con plantas que difieren en su morfología y estructura(8).
Al inicio de este documento se definen y describen los componentes del comportamiento de ingestión y las escalas espacio temporales del ambiente de pastoreo; luego, se presentan conceptos sobre las dimensiones del bocado, para analizar su relación con la densidad del forraje e impacto en la ingesta de forraje. Más adelante, se aborda la importancia de la altura de la pradera y su reducción, como medidas de abundancia de forraje, sobre el comportamiento de ingestión y el consumo. Al final, se describen el patrón de actividad de pastoreo de vacas y resultados de comportamiento de ingestión y consumo.
Comportamiento de ingestión y escalas de pastoreo
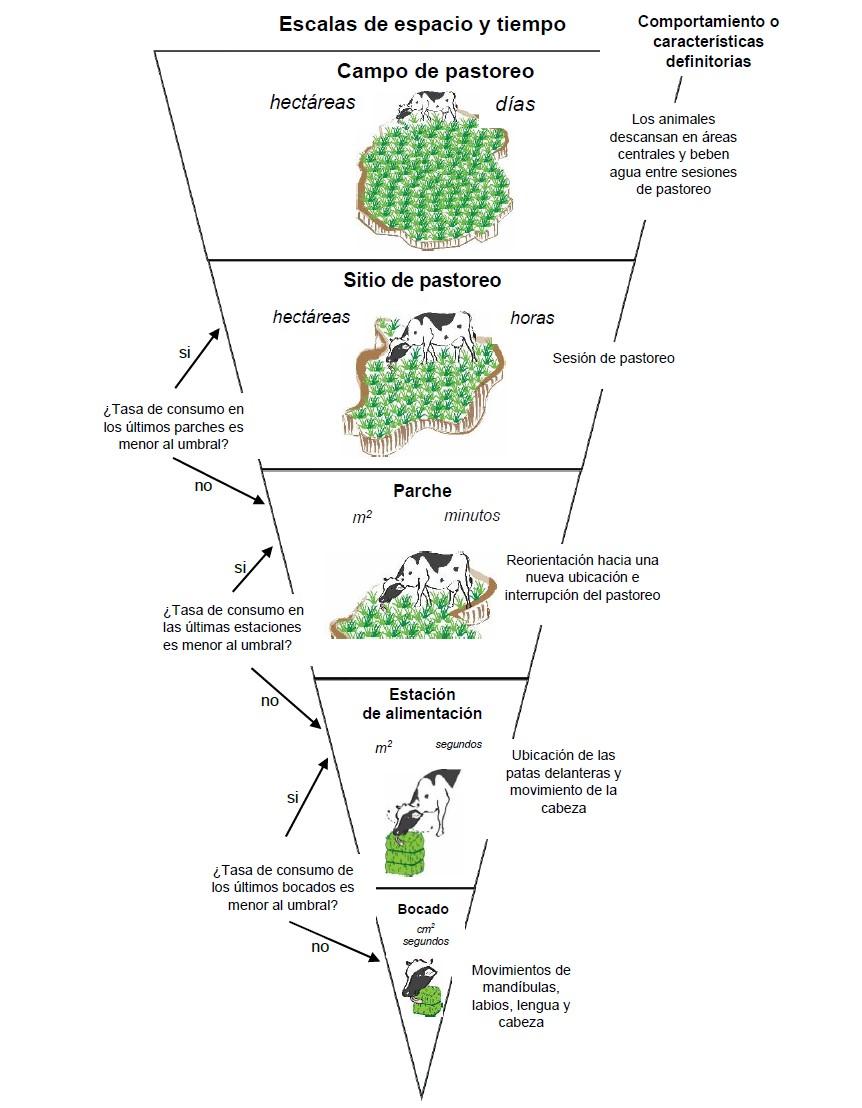
El conocimiento del CI de las vacas es imprescindible para comprender y gestionar su CF. En el análisis del CI de las vacas para evaluar el CF (g MS d-1), se integran componentes de la conducta de los animales y atributos de la pradera (Figura 1). Los componentes son MB (g MS bocado-1), tasa de bocados (TB, bocados min-1), TC (g MS min-1) y tiempo de pastoreo (TP, min d-1). Las vacas colectan el forraje en diferentes escalas jerárquicas de espacio y tiempo del ambiente de pastoreo, tomando decisiones, que en su conjunto se conocen como comportamiento de ingestión, equivalente a la dinámica de forrajeo (Figura 2)(9).
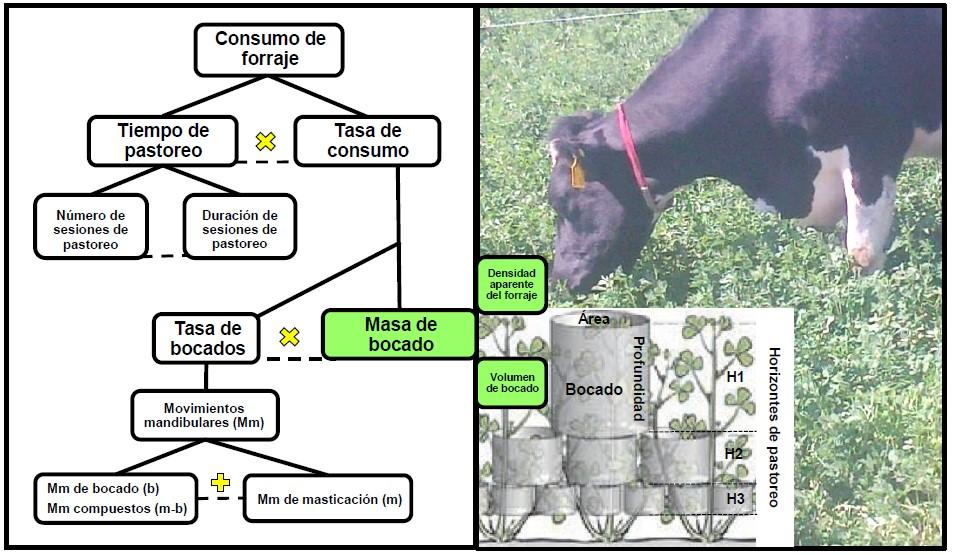
Figura 1: Componentes del comportamiento de ingestión de vacas en pastoreo(1,10,11).
La escala más pequeña de la dinámica de forrajeo es el bocado y su obtención se define como la colocación de forraje en la boca y su desprendimiento del resto de la planta por movimientos de la boca y de la cabeza(8,12). Con movimientos de los labios, lengua y mandíbulas ocurre la aprehensión y acomodo del bocado dentro de la boca y con movimientos de la cabeza se obtiene la tensión para lograr la ruptura del forraje. Durante esta operación, los bovinos utilizan la lengua para introducir el forraje en su boca y para ampliar el área de bocado(13).
Figura 2: Escalas espaciales y temporales del comportamiento de pastoreo de herbívoros grandes(9,11,12)
La escala espacial siguiente es la estación de alimentación, área en la que el animal selecciona y toma bocados con movimientos del cuello sin mover sus patas delanteras(14). El nivel inmediatamente superior es el parche, que consiste en un conjunto de estaciones de alimentación(13).
El forrajeo en estas escalas puede durar desde 1 seg hasta 30 min, en superficies que van desde pocos cm² hasta 1 ha(9). La escala siguiente es el sitio de pastoreo, que está integrado por distintos parches donde la vaca consume forraje durante una sesión de pastoreo(13). El tiempo de permanencia en esta escala varía entre 1 y 4 h, en superficies que van desde 1 a 4 ha(9). Por último, la escala superior del comportamiento de pastoreo es el campo de pastoreo, que implica mayor tiempo (semanas) y espacio (km²). Dicha escala puede ser alcanzada por hatos de ganado que, durante el pastoreo, se mueven dentro de grandes pastizales(15). El tiempo de permanencia de los grandes herbívoros (incluidos los bovinos) en las diferentes escalas, en general es función de la cantidad y calidad del forraje que allí se encuentra(9); además de otros factores que incluyen la topografía del área(16), la ubicación del agua(17), la estación del año(18), así como componentes del comportamiento social del ganado(19). El genotipo del animal es un factor en el que se han encontrado resultados contradictorios, en una investigación se comparó el pastoreo de vacas Beefmaster × Simford con el de vacas Baladi de menor tamaño, las vacas Baladí presentaron mayor tiempo de pastoreo y distancia recorrida(18). En otro estudio, compararon toretes de maduración temprana (Angus y Hereford) y maduración tardía (Limousin, Charolais) y no encontraron diferencias en comportamiento ingestivo y consumo entre genotipos(20). Sin embargo, el consumo de forraje en vacas F1 Hereford × Angus durante la gestación fue menor que en vacas Hereford y Angus(21).
En la investigación sobre el CF por vacas lecheras en praderas, han predominado los estudios de corto plazo, en las escalas de bocado(22,23), estación de alimentación(24,25), parche(26,27) y sitio de pastoreo(16). Una causa de que la investigación se haya focalizado más en el corto plazo es porque pocos investigadores cuentan con acceso a la grabación automática del CI y personal capacitado para hacer observaciones a largo plazo(7). Sin embargo, la ganadería de precisión, que abarca el uso de herramientas tecnológicas para evaluar el comportamiento del ganado(28), puede contribuir a tomar decisiones sobre la gestión en tiempo real y comprender mejor la relación planta-animal en los sistemas de pastoreo. Al respecto, existen diversos sensores y dispositivos que se han utilizado para el monitoreo de movimientos mandibulares y el comportamiento de ingestión de bovinos en pastoreo(12).
El CF de los animales en pastoreo se puede describir aritméticamente como el producto de dos componentes, TC y TP(1) y, a su vez la TC como el producto de TB y MB(29). Por su parte, la TB está determinada por los movimientos mandibulares (Figura 1) que pueden ser diferenciados con equipos que registran ya sea señales mecánicas(30) o acústicas(31). Los movimientos mandibulares son de masticación, bocado y compuestos (masticación-bocado)(23), estos últimos han sido identificados exclusivamente con el análisis acústico(32). Un metaanálisis de publicación reciente(6) que involucró 103 publicaciones con 278 experimentos, confirmó que la MB es un componente fundamental del CI en pastoreo, ya que es sensible a las principales características del dosel de la pradera y es factor determinante para la TC y el CF.
Para comprender la estrategia de forrajeo de los animales en pastoreo y poder optimizar el CF entre las escalas, hace más de dos décadas se postuló en un modelo que el desplazamiento entre las escalas más pequeñas está gobernado por la TC de las escalas jerárquicas inmediatamente inferiores (Figura 2)(33). Por ejemplo, cuando una vaca cosecha bocados dentro de una estación de alimentación, el pastoreo puede continuar hasta que la TC de los últimos bocados cae por debajo de un umbral. Si esa situación se presenta, la vaca se desplazará hacia la próxima escala jerárquica superior, en la que se realizará la cosecha de bocados a nivel de parche, y ahí a su vez permanecerá hasta que la TC llegue al umbral inferior.
Al respecto, se ha destacado que el tiempo de permanencia de animales cosechando bocados en cada estación de alimentación refleja la condición del dosel forrajero; cuando la calidad estructural del forraje es mejor en relación hoja/tallo, masa, altura y densidad de forraje mayor será el tiempo de permanencia(13). En un estudio se documentó que bovinos y ovinos permanecieron menos entre estaciones de alimentación y se desplazaron más rápidamente entre ellas en praderas nativas del sur de Brasil de baja altura (4 y 8 cm), que de mayor altura (12 y 16 cm)(14). Asimismo, en otro estudio se obtuvo que el tiempo de permanencia de novillos en la estación de alimentación de ballico anual (Lolium multiflorum Lam.) con avena negra (Avena strigosa Schreb) aumentó con incrementos en la altura de la pradera (10, 20, 30 y 40 cm)(34).
En relación con el CF a nivel de estación de alimentación, en una investigación se obtuvo que novillos pastoreando trigo (Triticum aestivum L.) de mayor altura (23.6 cm) tuvieron 1.9 veces mayor TC de forraje y cosecharon más bocados en el área pastoreada por estación de alimentación, que en praderas de menor altura (20.4 y 19.5 cm)(35). Esto es evidencia de que la estrategia de forrajeo que emplean los bovinos para lograr una alta ingesta de forraje durante el pastoreo, es aumentar su TC en parches con alta cantidad de forraje (mayor altura) y desplazarse más rápidamente entre estaciones de alimentación cuando se encuentran con parches de menor oferta de forraje (menor altura).
La distancia entre los parches potenciales para la ingesta de forraje también es importante en los componentes de la TC. Al respecto, se obtuvo que el número de bocados, los tiempos de permanencia, la velocidad de desplazamiento y la proporción de forraje total consumido por vacas en parches de alfalfa (Medicago sativa L.) y festuca (Festuca arundinacea L.) se incrementaron y la TC se redujo con el aumento en la distancia entre parches (1, 4 y 8 m)(27); las vacas realizaron un uso más uniforme de las especies a medida que la distancia entre parches fue mayor.
Dimensiones del bocado
El CF de un animal en pastoreo está relacionado con la capacidad que tiene su aparato recolector(11) y con la dinámica de la respuesta funcional, que es la relación que hay entre la TC y algunas variables que describen la abundancia de forraje en el área de pastoreo, por ejemplo, biomasa, altura y densidad del forraje(36). Por ello, el CF por bocado se puede evaluar por las dimensiones del bocado y la densidad aparente del forraje (DAF). La MB se puede cuantificar con el uso de aritmética, como el producto del volumen del bocado (VB) y la DAF(1). El VB no es el volumen de la cavidad bucal, sino el que ocupaba el forraje en la pradera que es cosechado con el bocado, el cual se ha considerado un cilindro de profundidad y área determinadas (Figura 1). Como resultado de un metaanálisis, se publicó que hay una relación curvilínea entre el VB (y) y la altura de la pradera (x), y=9.63*(1-exp (0.00125x)), n=90, RMSE=0.30, debida a las respuestas que hay entre la profundidad y el área del bocado por efecto del aumento de la altura de la pradera(10).
La profundidad del bocado se puede definir como la diferencia entre la altura media inicial de los hijatos y la altura media de los hijatos medida después del pastoreo(8) y en praderas de gramíneas puede conocerse a partir de la medición del largo de los hijatos extendidos antes y después del pastoreo(14). Además, resultados obtenidos en diferentes especies de forrajes y rumiantes ponen de manifiesto que hay una relación lineal entre la profundidad del bocado (y) y el largo de los hijatos extendidos (x), y=1.1+0.52x, R2=0.84, n=203, lo que destaca que la profundidad de bocado corresponde al 52 % del largo de los hijatos(1). El valor de la pendiente del modelo anterior se aproxima a la pendiente obtenida con otro modelo basado en datos de experimentos realizados con bovinos, que indicaron que la profundidad del bocado (y) se incrementa de manera lineal con la altura de la pradera (x), y=1.41+0.44x, RMSE=1.4, n=149(10).
Con base en lo anterior, hay evidencia(1,10) para sustentar que la profundidad del bocado en bovinos se aproxima a 50 % de altura de la pradera. No obstante, en un experimento previo sobre profundidad de bocado(26), se concluyó que hay que tener precaución con este concepto de proporcionalidad, a pesar de que en el mismo experimento se documentaron valores de profundidad de bocado en vacas de 40 a 55 % de altura de una pradera de ballico inglés (Lolium perenne L.). También, se reportó que la profundidad del bocado aumentó de manera progresiva durante los primeros 10 a 20 bocados, lo que indica que el ganado puede ser cauteloso al evaluar los parches durante el pastoreo(26).
El área del bocado en estudios con parcelas construidas manualmente se calcula como el cociente del área total pastoreada entre el número de bocados realizados(37). El área del bocado en bovinos aumenta con la altura de la pradera, con una relación curvilínea y un máximo teórico de 153.6 cm2(10). La DAF se refiere a la relación entre la masa del forraje y el volumen que ocupaba dicha masa en la pradera. En estudios sobre dimensiones del bocado la DAF se estima por el cociente de la masa de forraje y el volumen del estrato del dosel(38). La disminución de la DAF tiene un efecto positivo en el área de bocado de bovinos; en valores más bajos DAF el área de bocado fue cercana a 130 cm², mientras que para los valores altos de DAF el área de bocado presentó un valor mínimo de 33 cm²(10).
La altura y densidad de las praderas templadas no varían de manera independiente, sino que por lo regular están inversamente relacionadas. En praderas muy cortas (en general más densas) no son posibles mayores áreas de bocado, ya que los componentes cortos (esencialmente hojas) que se ubican en los bordes del área del bocado que el animal intenta tomar, escapan a la captura por la lengua y la prensión por medio de los dientes y el rodete dental(12). Por el contrario, en las praderas de clima templado de mayor altura en estado vegetativo la profundidad del bocado y el área de bocado son mayores debido a que se facilita la aprehensión de los componentes del dosel(13). En estos casos los bovinos ejecutan movimientos mandibulares compuestos; el animal toma un nuevo bocado cuando aún se encuentra masticando el bocado que tomó previamente(23).
La altura de la pradera en el consumo
Hasta ahora se han destacado los efectos de la altura de la pradera y la DAF en las dimensiones del bocado. Pero es importante discutir el impacto que tiene la altura de la pradera sobre el CF en pastoreo y, explicar cómo este último está mediado por los componentes del CI y por la MB(6,10). Sin embargo, en praderas templadas y tropicales, en estado vegetativo, el CF será afectado de manera positiva por la altura de la pradera, siempre y cuando se mantengan doseles con alta densidad de hojas verdes(29,39,40).
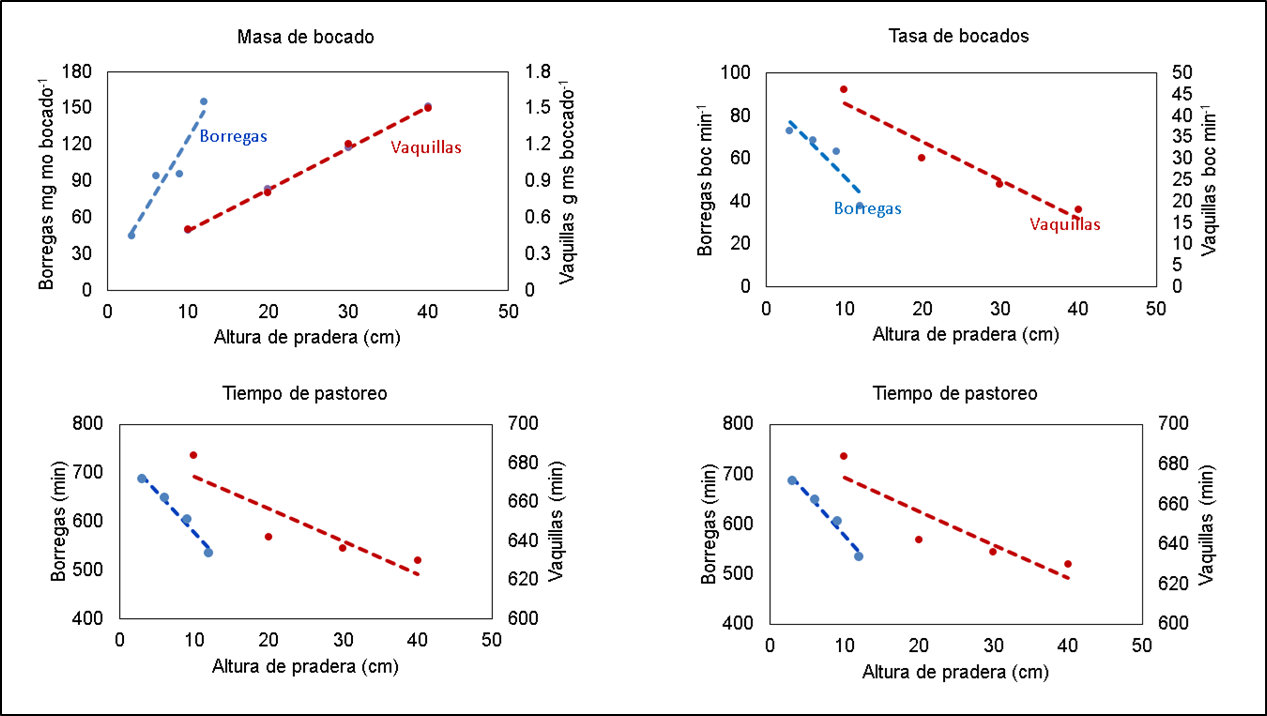
El aumento del CF por vacas lecheras en pastoreo es una respuesta clásica al incremento en la altura de pradera, la que se explica por los cambios que ocurren en los componentes del CI durante el pastoreo, también por efecto de cambios en la altura de pradera (Figura 3). Esto se evidenció desde un trabajo clásico realizado con borregas en praderas templadas(40) y en los últimos años, resultados obtenidos con bovinos de carne en praderas tropicales(29) confirmaron lo mismo. La respuesta clásica de la TC y la MB por efecto de la altura de pradera es que dichas variables se incrementan con el aumento de la altura(29,40,41,42). No obstante, estudios recientes en los que se evaluaron diferentes alturas de pradera en Cynodon sp. (10, 15, 20, 25, 30 y 35 cm) y Avena strigosa Schreb (15, 20, 25, 30, 35, 40, 45 y 50 cm), destacaron que si bien se encontró el mismo patrón, la TC y la MB disminuyeron en las praderas más altas que se evaluaron(24); los resultados evidenciaron una respuesta funcional en forma de cúpula, donde la TC de forraje fue mayor en alturas de pradera intermedias en ambas especies (39.2 g MS min-1 con 19 cm en Cynodon y 54 g MS min-1 con 29.3 cm en avena)(43). Los autores del trabajo resaltaron que dicha respuesta fue el resultado de cambios en la MB en las praderas altas (30 y 35 cm en Cynodon, 45 y 50 cm en avena) y atribuyeron la disminución de la MB a una reducción en su volumen, debida a menor área de bocado impulsada por el comportamiento selectivo de los animales, ya que no se relacionó con restricciones de la pradera que limitaran la formación del bocado(43).
Con el aumento de la altura de pradera, los bovinos también incrementan el total de movimientos mandibulares por gramo de MS consumida(38), porque los animales tienen que ejecutar mayor número de movimientos mandibulares por cada bocado debido a que estos son de mayor masa y, por ello, requieren mayor número de masticaciones para que se pueda tragar el forraje(6,41). En resumen, la MB y el CF en praderas templadas, en estado vegetativo, pueden resultar mayores en praderas con alturas intermedias(43) y altas(29), y para procesar bocados de mayor tamaño, los bovinos incrementan el número de movimientos mandibulares compuestos(23).
Otro cambio importante en el CI de las vacas en pastoreo en respuesta al incremento en altura de pradera es la reducción en la TB y TP (Figura 3); la primera es consecuencia de la mayor cantidad de forraje aprehendido por bocado, que implica un aumento en el tiempo por bocado(24) por el incremento del número de movimientos mandibulares de masticación(34). En otras palabras, a medida que los bovinos aprehenden más forraje por bocado emplean más tiempo para masticar, lo que aplaza la toma del siguiente bocado(6,34).
La reducción de la altura de la pradera en el consumo
Hasta ahora se ha referido cómo el CF se explica por el CI de los animales y por características estructurales del dosel de la pradera (cantidad de forraje, DAF y altura de pradera) que regulan la MB, con base en estudios donde se evaluaron dichas variables en plantas en estado vegetativo de diferentes alturas. Sin embargo, es importante discutir el efecto que, sobre el CF tiene la reducción de altura de la pradera que se presenta con el agotamiento rápido del recurso forrajero que ocurre durante el pastoreo a través del día. La cosecha de bocados durante el pastoreo se realiza a través de horizontes del dosel forrajero, a una profundidad más o menos constante de 50 % de altura de la pradera(11).
Los bovinos inician el pastoreo en el horizonte 1 (H1), consumen 50 % de la altura del forraje y después continúan al horizonte 2 (H2), en el que cosecharán de manera aproximada la misma proporción de altura correspondiente al horizonte, hasta llegar al horizonte 3 (H3) y finalizar a una altura límite de pastoreo (Figura 1). En una investigación realizada con bovinos se obtuvo que la profundidad, el área y la MB en trigo, sorgo (Sorghum sacharatum L.) y alfalfa (Medicago sativa L.) variaron a través de los horizontes de pastoreo(22). Desde el H1 hasta el H3, las profundidades de bocado disminuyeron en 76, 78 y 70 %, las áreas de bocados se redujeron en 44, 56 y 56 % y en consecuencia las MB fueron menores en 61, 71 y 87 %, respectivamente para trigo, sorgo y alfalfa.
En la estructura vertical de la pradera se encuentra parte de la causa de la variación en las dimensiones del bocado, porque el aumento del seudotallo, tallo y material muerto desde la superficie de la pradera hasta su base, se vuelven barreras para la formación del bocado(11,38), y en consecuencia modifican la MB y a su vez el CF. También, se ha documentado que el seudotallo y las alturas de rebrote y material muerto dentro del dosel forrajero, son reguladores parciales de la profundidad de bocado(26). Por lo anterior, la disminución del área de bocado desde H1 a H3 se debe a la disminución de hojas en la base del dosel forrajero y a la dificultad de aprehensión del forraje que escapa al barrido de la lengua(22). Además, el aumento de la DAF en los estratos más bajos de las praderas(38,39), afecta de manera negativa el área de bocado(10).
Con base en lo anterior, hay evidencia para destacar que la disminución del CF que se presenta con la reducción de la altura de la pradera, cuando los animales son forzados a cosechar hasta el horizonte más bajo de la pradera, se debe a la cosecha de bocados pequeños. El pastoreo en franjas se asocia con situaciones de rápido agotamiento del recurso forrajero y cuando los niveles de reducción de altura de pradera en dicha situación no se controlan, el CF puede ser afectado de manera negativa(44). Sin embargo, en situaciones de gestión diferente, en las que las praderas no presentan cambios en su condición durante el día, la TC es constante y el comportamiento del consumo es similar a través del día(29). El pastoreo rotatinuo (baja intensidad y altas frecuencias de pastoreo) es una estrategia de gestión para mantener altas TC de forraje, ya que los animales cosechan la mayoría de los bocados en el H1(1,45).
Por tanto, es importante resaltar que el control de la intensidad del pastoreo durante la gestión de la pradera es una medida de manejo útil para tener una idea del nivel de CF. Por ejemplo, para maximizar el CF en vacas lecheras en pastoreo de Lolium arundinaceum, se ha destacado que la gestión del pastoreo debe realizarse a bajas intensidades de pastoreo; alturas de forraje residual de 12 y 15 cm en otoño-invierno y primavera, respectivamente(25). Mientras que, en bovinos bajo pastoreo de avena negra cv. Iapar 61, Cynodon sp. cv. Tifton 85(24) y sorgo (Sorghum bicolor L.)(46) se encontró que altas TC se mantenían hasta un nivel de reducción de 40 % de altura de la pradera. El forraje residual más corto, no solamente se traduce en menor consumo, sino que también en menor selectividad lo que contribuye a menor ingesta de materia orgánica digestible, y por tanto menor producción de leche(25).
Los bovinos y ovinos en pastoreo son capaces de aumentar su TB y TP ante situaciones de baja MB (Figura 3) como estrategia conductual para compensar de manera parcial la reducción en la TC(6,7). No obstante, no se compensa de manera completa la disminución de la cantidad de forraje ingerido por bocado y como resultado el CF es menor(7). Por diferencias en conducta animal, la respuesta se presenta en pastoreo continuo y rotacional pero no así en pastoreo en franjas; en estos casos, la altura se torna en factor muy limitante, ocasionando que los animales opten por no pastorear durante más tiempo por la baja retribución asociada a masas de bocados muy pequeñas(20,47).
El tiempo de pastoreo y el consumo de forraje
Durante el día las vacas realizan actividades de pastoreo, rumia y otras acciones. Algunos resultados del tiempo que dedican las vacas a cada actividad en clima templado en pastoreo de ballico inglés se presentan en el Cuadro 1. Las vacas en praderas templadas exhiben diferentes sesiones de pastoreo durante el día, entre tres y cuatro sesiones, dos de mayor intensidad; al medio día y antes del anochecer(2). En un estudio en pastoreo de ballico inglés con asignación de forraje por la mañana, vacas invirtieron entre 70 y 80 % de su tiempo en pastoreo después de las ordeñas matutina y vespertina(48). Al respecto, en otra investigación(49) se encontró que conforme transcurrieron las primeras 4 h después de las ordeñas matutina (después de las 0800 h) y vespertina (después de las 1500 h) el porcentaje de vacas en pastoreo declinó de 94 a 35 % (ordeña matutina) y 87 a 9 % (ordeña vespertina).
En relación con los tiempos de rumia e inactividad por vacas lecheras, éstas dedican más del 70 % de su tiempo a dichas actividades por la noche(48,49). A pesar de que el mayor tiempo de rumia se presenta durante la noche, también hay periodos de rumia en el día. La rumia durante la noche se asocia con la conducta natural que exhiben los rumiantes; al atardecer consumen forraje lo más rápido posible y reservan la rumia para la noche, cuando se ocultan con relativa seguridad y disminuyen los riesgos de ser depredados(2).
Debido a que el tiempo total de pastoreo diario es el resultado acumulado de todos las sesiones o eventos de pastoreo, con fines de estimación de CF el TP debe ser de pastoreo activo, el cual está determinado por el número y la duración de las sesiones de pastoreo (SP) durante el día(13). La SP por definición se refiere a una secuencia larga de pastoreo, la cual se caracteriza por un mínimo de 20 min de pastoreo activo, cuya interrupción ocurre por la realización de cualquier otra actividad, también por un periodo mínimo de 20 min(50).
A partir de investigaciones realizadas en pastoreo rotacional de praderas dominadas por ballico inglés, se ha encontrado que vacas lecheras pueden efectuar alrededor de 5.6 a 10.0 SP a través del día (Cuadro 1) y la duración de la SP es de aproximadamente 90 min. El número de SP y su duración se han asociado con la calidad y cantidad de forraje; si la masa de forraje disponible es elevada, el número de SP es mayor y su duración es menor(25). En esas situaciones los bovinos se vuelven más selectivos y por ende pueden cosechar forraje de mayor calidad en menor tiempo(50).
El CF en vacas lecheras bajo pastoreo de praderas dominadas por ballico inglés es de aproximadamente 3 % de su peso vivo (PV), a partir de la integración de resultados sobre componentes de CI reportados(48,51) (Cuadro 1). No obstante, la variación alrededor de esa media puede ser elevada, debido a factores de la pradera, de gestión del pastoreo y del animal. En una revisión sobre pastoreo de praderas templadas, se reportaron valores de CF en vacas lecheras de 1.6 a 3 % del PV.
Tasa de consumo de diferentes categorías
Con base en los resultados del Cuadro 1, la TB es similar entre vaquillas y vacas adultas (en promedio 59 bocados min-1), en cambio, la MB promedio es mayor en animales adultos (0.41 g) que en jóvenes (0.22 g). Lo anterior se debe a la mayor longitud en la arcada de los incisivos de las vacas, porque determina el área y el volumen de bocado y en consecuencia la MB(10). Por lo anterior, la TC promedio reportada en las vaquillas es menor (12.8 g MS min-1) que en las vacas (24 g MS min-1).
Conclusiones
Aunque las vacas en praderas de baja altura (cortas) tienen la capacidad de aumentar la TB y el TP, como estrategia conductual para remediar la reducción en la TC debida a la cosecha de bocados de menor peso, los animales no compensan la disminución de la cantidad de forraje ingerido por bocado y como resultado el CF puede ser bajo. Por tanto, cuando las vacas se mantienen en praderas muy cortas o en situaciones de sobrepastoreo es seguro que las TC serán bajas y en consecuencia el consumo diario de forraje será menor. Debido a la relación que tiene la altura de la pradera con los componentes del CI y el CF, en la práctica el monitoreo de la altura de la pradera puede ser una herramienta útil para evaluar el CF y mantener altas TC en los bovinos. No obstante, en México hace falta desarrollar investigaciones enfocadas a evaluar la TC de bovinos en pastoreo a diferentes alturas de pradera, que permitan generar implicaciones prácticas para la gestión del pastoreo.
Agradecimientos y conflicto de interés
Los autores agradecen el apoyo del CONACYT para los estudios de doctorado del primer autor y declaran que no tienen ningún conflicto de interés.
Literatura citada:
Figura 3: Consumo de forraje y componentes del comportamiento de ingestión en respuesta a la altura de pradera(29,40)
Cuadro 1: Componentes de la conducta de ingestión, tiempos de pastoreo, rumia e inactividad en vacas lecheras bajo pastoreo en praderas de clima templado
Vacas lecheras | Vaquillas | |||
Ballico inglés(52) | Ballico inglés(53) | Ballico inglés(51) | Trifolium repens(51) | |
Consumo de forraje, kg MS 100 kg PV-1 | 3.3 | 3.0 | 3.0 | 2.4 |
Consumo de forraje, kg MS día-1 | 15.2 | 15.5 | 6.9 | 5.5 |
Tiempo de pastoreo, min día-1 | 629 | 646 | 536 | 436 |
Tasa de consumo, g MS min-1 | 24.2 | 23.9 | 12.8 | 12.7 |
Tasa de bocados, bocados min-1 | 62 | 57 | 61 | 55 |
Masa de bocado, g | 0.39 | 0.42 | 0.21 | 0.23 |
Tiempos de pastoreo, rumia y otras actividades en vacas lecheras | ||||
Ballico inglés(48) | Ballico (80 %) y Trébol(48) | Ballico inglés(53) | Ballico (80 %) y Trébol(54) | |
Tiempo de pastoreo, min día-1 | 622 | 591 | 646 | 549 |
Pastoreo, sesiones día-1 | 6.5 | 5.6 | 10.0 | 9.6 |
Duración de sesión, min período-1 | 99 | 120 | 79.5 | 63 |
Rumia, min día-1 | 431 | 402 | 426 | 401 |
Otras actividades, min día-1 | 388 | 447 | 368 | 490 |