https://doi.org/10.22319/rmcp.v15i2.6090
Review
Implication of Fusariotoxins in poultry production. Review
Gabriela Guadalupe Gómez Verduzco a
Ernesto Ávila González a
Guillermo Téllez Isaías b
Juan Carlos Del Río García c
Jacqueline Uribe Rivera c*
a Universidad Nacional Autónoma de México. Facultad de Medicina Veterinaria y Zootecnia. Ciudad de México, México.
b Universidad de Arkansas. Departamento de Ciencia Avícola. Arkansas, Estados Unidos de América.
c Universidad Nacional Autónoma de México. Facultad de Estudios Superiores Cuautitlán. Carretera Cuautitlán-Teoloyucan Km. 2.5, San Sebastián Xhala, CP 54714 Cuautitlán Izcalli, Estado de México, México.
* Corresponding author: juribe_mvz@hotmail.com
Abstract:
Mycotoxins are secondary metabolites produced by fungi of various genera. Among the most important mycotoxins are those produced by fungi of the genus Fusarium sp., which can be divided into several groups for study, which are the groups of trichothecenes (and T-2 toxin), fumonisins, mainly fumonisin B1 (B1, B2, B3, B4, A1, and A2), and zearalenone with estrogenic effects. Although fusariotoxins cause similar effects because they share the same mechanism of action, by altering protein synthesis in intoxicated poultry, it is important to mention the incidence as well as the characteristics between each of them. Therefore, the characteristics of each group mentioned are described in each section.
Keywords: Mycotoxins, Fusariotoxins, Poultry, Fungi.
Received: 03/11/2021
Accepted: 31/01/2024
Introduction
Within livestock production, feed accounts for 65 to 70 % of the total cost of production; however, despite constant efforts to achieve the safety of food intended for animal production, there are still factors that diminish its quality, such as biological pathogens (such as Salmonella spp., E. coli, Listeria spp., Campylobacter spp.), chemical substances (fungicides, herbicides, and insecticides), and presence of fungi (Fusarium, Penicillium, Mucor etc.)(1). Fungal contamination can occur under various conditions associated with both environmental factors and factors associated with the fungus, such as size and species(2).
Mycotoxins are secondary metabolites produced by filamentous fungi(3), considered toxic substances that present themselves as organic compounds of low molecular weight, which is why they do not have immunogenicity characteristics. The production of mycotoxins depends on a series of environmental factors such as humidity, temperature, ventilation, constitution of the substrate in which the fungus develops, damage to the integrity of the grains, and interaction between the various fungi present in the substrates(4). It is mentioned that 25 % of crops worldwide are contaminated with mycotoxins(5).
Fusariotoxins
Among the toxins reported with the highest incidence within animal production are those produced by different species of fungi of the genus Fusarium sp, which are capable of inducing acute effects, as well as chronic effects, depending on the type of mycotoxin, the level and duration of exposure, and the species and age of the animal.
Avian species are considered resistant to the occurrence of fusariotoxin intoxication, which has been explained either by low sensitivity to toxicity mechanisms or by differences in toxicokinetic properties(6). These fungal species have been classified within the group of field fungi, with grains requiring a high percentage of moisture (approximately 20 to 22 %) for the production of mycotoxins. The main substrate where the production of fusariotoxins is observed is corn; nevertheless, their growth has also been reported in other substrates such as sorghum, wheat, oats, barley, and soybeans. Fusariotoxins, in turn, are classified into three groups for study, among which are the trichothecenes group (which in turn is classified according to the presence or absence of a macrocyclic ring in their chemical structure), the zearalenone group and the fumonisin group (Table 1).
Table 1: Mycotoxins produced by fungi of the genus Fusarium sp.
Adapted from(11,94,95).
Experimental studies have agreed that the main effect of the toxins produced by Fusarium sp. has a direct impact on intestinal integrity (increased intestinal permeability), an effect correlated in turn with a decrease in the animal’s immune response (alteration in the production of cells involved in the immune response)(7,8). Both effects have their origin in the alteration of protein synthesis that takes place in the different metabolic processes in the animal(9,10). It is important to mention that regardless of the mycotoxin or mycotoxins that are present in the feed, the cells of the digestive tract are the first to be in contact with the mycotoxins; this means that, possibly, the entire intestinal epithelium can be compromised even before the absorption of the feed begins, and in turn, it can also be affected by mycotoxins that have a low absorption rate, such as fumonisins and deoxynivalenol. Although fusariotoxins may have similar mechanisms of action among them, it is important to mention the individual characteristics of each(11).
Trichothecenes generalities
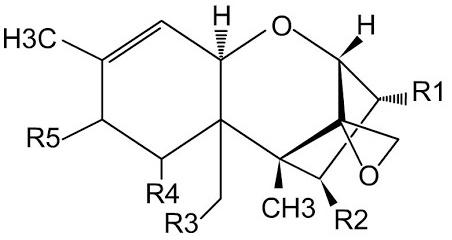
More than 150 different types of trichothecenes have been identified, and it has been established that, based on their occurrence in food, the most prevalent are T-2 toxin, deoxynivalenol (DON or vomitoxin), and diacetoxyscirpenol (anguidine), which in their chemical structure consist of a tetracyclic sesquiterpene skeleton, an oxane ring, and a stable epoxide group (12,13-epoxytrichothecene) that confers its toxicity (Table 2)(7). The main substrates where can be find the presence of trichothecenes are mainly corn, wheat, barley, oats, rice, and soybeans. Broadly speaking, trichothecenes are considered to be cytotoxic, immunosuppressive and inhibitors of protein synthesis(8). Although trichothecenes can generally cause gastrointestinal, dermatotoxic, immunotoxic, and genotoxic effects, it has been reported that, under acute intoxication, they can be identified by evident skin inflammation, diarrhea, edema, dermal necrosis, hemorrhages in the mucosa of the gastrointestinal tract, and negative alterations in productive parameters(12,13,14).
Table 2: Structural differences in trichothecenes present in poultry production
Adapted from(18).
T-2 toxin
Main characteristics and chemical structure
The T-2 toxin, belonging to group “A” of trichothecenes, contains in its tetracyclic chemical structure a sesquiterpenoid system and an epoxy-trichothecene group in C-12 and C-13. It can mainly be found in corn, wheat, oats, and barley(15,16).
It has been determined that the main fungus producing this mycotoxin is Fusarium tricinctum, however, it can be produced in turn by Fusarium acuminatum, nivale, oxysporum, poae, solani and sporotrichioides. In relation to this, the environmental conditions required for the production of the fungus are an average environmental temperature of 18 to 30° C, as well as a relative humidity of around 95 % and a water activity (AW) value greater than 0.88 (which must be 0.91 for the fungus to produce the mycotoxin)(17). The chemical structure of this mycotoxin makes it a non-volatile compound soluble in acetone, chloroform and ethyl alcohol; it is of low molecular weight (approximately 466.52 g/mol) and is described as a mycotoxin highly resistant to heat (200-210 °C) and UV radiation, therefore, it is not easily inactivated in stages during feed processing; nevertheless, it has been reported that the addition of sodium hypochlorite or hydroxide for a period of 4 h can work as an inactivation method(18).
It has been reported that the maximum levels allowed within the European regulation in finished feed for T2 toxin range from 0.2 to 2 mg/kg of finished feed(19,20) and a concentration of 4.97 mg/kg of feed has been established as LD501 and 10 mg/kg as LDPV2(18). It should be considered that T-2 toxin can generate interactions in the presence of other mycotoxins, which will be additive in the presence of deoxynivalenol (DON), ochratoxin A (OTA) and fumonisin B1 and synergistic in the presence of nivalenol and aflatoxins(21-25).
Metabolism
The main pathways by which T-2 toxin will be metabolized in poultry are mainly de-epoxidation and de-acetylation(26). In the case of de-epoxidation, which is the most common pathway, the result will be the loss of the “epoxy” group, which confers toxicity to the mycotoxin, while during the loss of the “acetyl” group, the result will be the obtaining of secondary metabolites of the toxin, such as HT-2 and T2-tetraol(6).
Mechanism of action and main effects in poultry
T-2 toxin and its derivatives base their toxicity on the negative alteration it will cause on protein, DNA and RNA synthesis, as well as on the cell cycle in different types of cells and its ability to induce apoptosis and necrosis, as well as lipid peroxidation, mainly in labile or actively producing cells. It interacts with the peptidyl transferase of the ribosomal 60s subunit, inhibiting the formation of new peptide bonds(27). Apoptosis caused by T-2 toxin was evidenced in cell lines such as Vero and human hepatocarcinogenic(28). In addition, it has been reported that T2 toxin reduces production parameters, with oral exposure to the toxin being the main route of access to the body(18).
Effects on the immune system
In general, T-2 toxin has a time/dose-dependent immunosuppressive effect, either high concentrations for a short period of time or low concentrations continuously(29). The presence of leukopenia has been reported, leading to an increased susceptibility to secondary infections (Listeria monocytogenes and Salmonella sp). T-2 toxin has also been associated with a decrease in the amount of antibodies against Newcastle disease and infection of the bursa of Fabricius(18). In addition, it has been described that it can act as an immunostimulant by increasing IgA levels; this is related to the abnormal and transient activation of genes involved in the inflammatory response(26,27). It is also mentioned that it can alter the maturation of antigen-presenting cells by modifying antibody levels by lymphocyte proliferation, leading to an increase in susceptibility to infectious agents(30).
Effects on the digestive system
Although T-2 toxin has a rapid absorption in the gastrointestinal tract and an elimination of approximately 80 to 90 %(19), its toxicity is mainly effected during the enterohepatic circulation, and the negative effect it has on the liver is the decrease of the enzymatic activity necessary for the metabolism of toxic substances and the induction of lipoperoxidation, which will consequently result in the formation of free radicals(18,31,32). It is important to mention that the alteration on mucous membranes will be an evident characteristic related to the diagnosis of mycotoxicosis caused by trichothecenes, especially by T-2 toxin, this related to the caustic effect of the toxin, and it will be visible particularly in the oral cavity, being observed as dermonecrotic lesions that will go from a whitish coloration at the beginning of exposure to a black coloration in chronic exposures. Necrotic lesions may also be observed in gizzards, intestinal mucosa, proventriculus and liver(33,34).
Effect on the nervous system and productive performance
The effect on the nervous system occurs due to the inhibition of protein synthesis, which leads to an increase in the concentration of the amino acid tryptophan, a precursor of serotonin, which in turn increases the concentration of serotonin, thus causing the activation of serotonergic neurons(18,35). The synergistic presence of DON with T-2 toxin can increase this effect, and anorexia, locomotor problems and vomiting can be observed(36,37); lesions present in the oral cavity should also be considered, which will contribute to the decrease in consumption, thus affecting body weight at the end of the cycle, also observing a deficient uniformity in the flock(19,38,39). In the case of laying hens, a decrease in egg production, as well as in shell quality and hatchability, can be observed(38).
Deoxynivalenol
Deoxynivalenol, also known as DON or vomitoxin, is a common fusariotoxin in production poultry, which may be more resistant than other consumption species(24,21,40). Deoxynivalenol can be found in finished products such as pasta, bread, cookies, and beer. Although it is considered a teratogenic mycotoxin, it has not been classified as carcinogenic, mutagenic, or genotoxic(41).
Characteristics and metabolism
DON is a stable organic polar compound with high resistance to acidic pH media and high temperatures (up to 180 °C). Its structure contains three free hydroxyl groups associated with its toxicity(42). The lower susceptibility of poultry has been related to their low percentage of intestinal absorption, which is approximately 5 to 20 %, while their excretion is 78.6 to 98.5 %, mainly through the bile duct, in a period of 24 to 72 h(43,44). In the case of poultry, the main metabolic pathways of DON are sulfation, glucuronidation, and de-epoxidation(45). When DON is metabolized through acetylation processes, compounds such as 3-acetyl-DON and 15-acetyl-DON will be obtained, which can also be found in feeds, and their importance lies in the fact that they can easily reverse DON and recover its toxicity(46). Other compounds derived from deoxynivalenol are DON-3-glucoside, as well as masked compounds that have gained importance for their ability to recirculate in the body, recovering their toxicity characteristic; in addition, they have been recognized as having the capacity to produce mixed toxic effects, causing adverse effects on animal welfare and productivity(15,47,48).
Inhibition of protein synthesis
Inhibition in protein synthesis occurs when DON forms a bond through the binding of its epoxide group with the ribosomal 60s fraction, causing an alteration in the binding of the latter with the 40s fraction, preventing the translation of messenger RNA and preventing the binding of amino acids to the polypeptide chain, either in elongation or during the termination of protein synthesis, also generating an increase in the amount of polyribosomes (80s), since the uncoupling of messenger RNA and the release of the peptide chain are inhibited(46). It has also been reported that it can alter the activity of the enzyme ribosomal peptidyl transferase by forming peptide bonds between amino acids(46,49). It is worth mentioning that changes in the conformation of ribosomes can induce a stress response, thus leading to the activation of mitogen-activated protein kinases (MAPKs)(50).
Effect on the immune system
An increase in serum IgA concentration has been observed, which has been related to the inhibition of protein synthesis of protein components necessary for the transport of IgA to the hepatobiliary system(11). Alterations in humoral immunity due to DON intoxication have been determined by evaluating the response to vaccination, mainly against Newcastle disease virus and infectious bronchitis virus, and a decrease in antibody titers has been observed. In the case of cellular immunity, induction of apoptosis in leukocyte cells such as T and B lymphocytes and macrophages, alteration of the activation of CD4 and CD8 T lymphocytes to CD4+ and CD8+, and a reduction in the serum concentration of TNF-α(51) have been observed. DON has the ability to alter the expression of genes that code for the production of pro-inflammatory cytokines and chemokines(52). This has also been related to the activation of genes that also code for the activation of cyclo-oxygenase-2 (COX-2) and nuclear factor Kappa of activated B cells (NF-ΚB)(26).
Effect on intestinal health
Deoxynivalenol is known to be a potent anorexic and emetic compound due to the alteration in the regulation of several signaling pathways. These alterations can affect the secretion of anorexigenic or orexigenic hormones, such as serotonin, released by enterochromaffin or Kulchitsky cells. It has also been observed that DON, by causing severe suppression of cytokine signaling processes, generates the activation of pro-inflammatory cytokines, affecting growth hormone signaling by suppression of two proteins, which are the hepatic acid-labile subunit as insulin-like growth factor and insulin-like growth factor I(52). In addition, an increase in transepithelial electrical resistance (TEER) in jejunum is mentioned, mainly attributed to the decrease in space between the tight junctions and a decrease in paracellular permeability to ions(53).
Effect of DON on productivity
In the case of laying hens, it has been reported that under a concentration of 2 to 3 mg DON/kg of feed, a slight decrease in egg production can be observed; however, fertility and hatchability can be maintained unchanged at this same concentration(45,46). It is important to mention that the effects may vary according to the dose ingested, and with the consumption of high concentrations (5 to 10 mg/kg of feed) of deoxynivalenol, diarrhea may be observed, preventing the animal from correctly assimilating the nutrients, delaying its growth, and causing unevenness in the flock(54-56). In the case of consumption of low concentrations, anorexia and stunted growth have been observed(6,46).
Fumonisins
Fumonisins are a group of toxins produced by several species of fungi (Table 1), within which the main producers of fumonisins are: Fusarium verticillioides and F. proliferatum(57). These fungi will be found as common contaminants of substrates such as corn, rice, sorghum, barley, peanuts, and cotton, where their optimal growth will be in temperatures between 22.2 and 27.5 °C with an AW of 0.97-0.98 for the production of the toxin(58-60).
Main features and metabolism
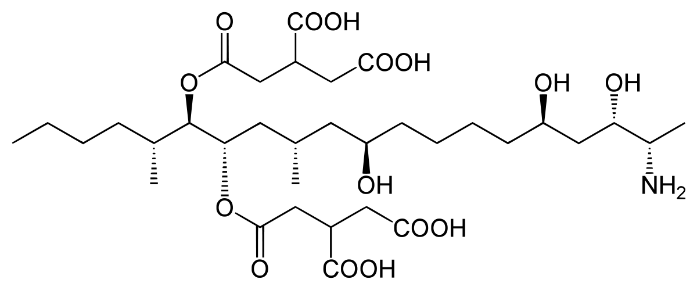
There are six types of fumonisins, B1, B2, B3, B4, A1 and A2 (Table 3); nevertheless, the most important due to their level of incidence and toxicity are B1 and B2. The basic structure of fumonisins consists of a 20-carbon alkylamine, with one or two hydroxyl groups and one or more methyl groups or esterified tricarballylic acid(61). Fumonisins are polar compounds that are soluble in water and organic compounds such as methanol and acetonitrile, but are insoluble in non-polar compounds, which in turn facilitates their elimination from the body(6). In the case of poultry, it is now known that, like pigs, acetylated or hydrolyzed derivatives (HFB1) are also generated as a product of acetylation after hydrolysis, as part of the toxicity mechanisms associated with their metabolism in the body(62-65).
Table 3: Basic chemical structure of fumonisins B1, B2, B3, and B4
Adapted from (66,96-98).
Mechanism of action of fumonisins
The mechanism of action of fumonisins is based on the interference in the metabolism of sphingolipids by competitive inhibition with sphinganine from de novo synthesis and sphingosine from sphingolipid turnover, generating an accumulation of sphingoid bases, blocking the synthesis of complex sphingolipids through the inhibition of the enzyme ceramide synthase(15,66,67). In addition, sphingosine and sphinganine have pro-apoptotic, cytotoxic, and growth-inhibiting effects(68). Sphingolipids can act as first and second messengers in a variety of signaling pathways and play a vital role in the formation of membrane microdomains called lipid rafts(69). Ceramide is involved in the processes of cell differentiation, senescence, and death. Sphingosine 1-phosphate (S1-P), on the other hand, promotes cell survival and proliferation(66,67). The immediate consequence of the inhibition of ceramide synthase is the accumulation of the sphingoid base that functions as a substrate, (Sa) sphinganine and, to a lesser extent, (So) sphingosine(66). The liver and kidneys are the main target organs, although variations have been observed depending on the species, dose, and sex(70-74). The gastrointestinal tract can also be a target organ for fumonisins since glycosphingolipids bind to sites for microbial pathogens and their toxins, through the inhibition of ceramide synthase in the digestive tract, it can alter the expression of glycosphingolipid binding sites or the transport of microbial toxins, and consequently the sensitivity of animals to infectious agents(17).
The different effects of fumonisins on different productive species have been reported, and it has been observed that they can vary from alterations in productive variables to changes in biochemical parameters and immune response.
Effect on production parameters
Although the negative effect on productivity has generally been observed in high concentrations, it has also been reported that, in concentrations around 5 ppm, it can cause low uniformity in production parameters, mainly on body weight at the end of the cycle in broilers(75).
Morphological and blood biochemistry alterations
The morphological alterations that have been observed in the case of production poultry are a decrease or increase in relative weight in organs (heart, liver, spleen, bursa of Fabricius, proventriculus)(76-78), hydropericardium, fatty liver or friable liver, hyperplasia of bile ducts, cardiac degeneration and necrosis, and loss of tonicity in gizzard and proventriculus(77,79). In addition, an increase in serum calcium and cholesterol values and a decrease in liver enzyme values (aspartate amino transferase, alanine amino transferase, lactate dehydrogenase, γ glutamyl transferase) have been observed, suggesting an injury to hepatic metabolism(80,81).
Effect on immune response
Among the lesions that have been observed as part of the damage of fumonisins to the immune system in poultry, hemorrhages, leukocyte infiltrations, fat infiltration, necrotic lesions, fibrosis in the liver, kidneys, lung, heart, intestine, gizzard, bursa of Fabricius and pancreas, as well as edema and hemorrhages in the brain are described. Cortical atrophy in the thymus, multifocal hepatic necrosis, and biliary hyperplasia have also been observed, leading to lymphoid depletion(82). On the other hand, a reduction in the size of the spleen may occur along with depletion of white pulp, thinning of cardiac myocytes, lymphoid cell depletion in the bursa follicles, and renal tubular nephrosis at dosages exceeding 150 mg/kg of feed(83-85). Studies on the effect of fumonisins have been conducted using high concentrations of fumonisin; however, it has been reported that, on average, a concentration of between 3 and 5 mg/kg of fumonisin B1 has been found in the main components of diets intended for animal feed, such as corn(86).
Zearalenone
Zearalenone (previously known as F-2 toxin) is a non-steroidal estrogenic (mainly in pigs), hematotoxic and genotoxic (rodents) fusariotoxin. The main fungi that produce zearalenone are F. graminearum, F. oxysporum, F. roseum, which require temperatures of between 21 and 25 °C and an AW of approximately 0.87 for the production of the toxin. In particular, it can be found contaminating corn, barley, rice, and soybeans, as well as finished products such as meals and beer(87). In the case of poultry, it is considered that the consumption of a high concentration is required to observe negative effects on production; in any case, it has been reported that turkeys and ducks are the most susceptible species in poultry, as is the case with other mycotoxins(88).
Metabolism and chemical structure
The characteristics of zearalenone allow it to be a compound of rapid biotransformation and excretion. This means that it is not easy to find zearalenone in poultry products(89). Zearalenone has a percentage of absorption by the digestive tract of approximately 10 % in poultry, and a high percentage of elimination of both zearalenone and its conjugation metabolites (approximately 65 %)(90,91). The structure of zearalenone is composed of a resorcyclic lactone ring like its main derivative, zearalenol (α-zearalenol), which will be a derivative with a higher degree of toxicity than zearalenone. In the case of poultry, the main pathways involved in zearalenone metabolism are sulfation and glucuronidation(6).
Mechanism of action
The main mechanism of action used by this mycotoxin is the disruption of endocrine metabolisms, this is done through interaction with nuclear estrogen receptors (ERs), with a direct effect on transcription (which is estrogen dependent) in the nucleus (mechanisms of competition with the same estrogens); although the structure of zearalenone is not similar to that of estrogens, it can place an OH group belonging to its lactone ring in an advantageous position for its interaction with estrogen receptors(92).
Alterations caused by zearalenone in poultry
In the case of roosters, a reduction in the size of the testicles can be observed, which can microscopically show fatty degeneration and atrophy of the germinal epithelium. Finally, production parameters can also be observed to be decreased, related to a decrease in feed consumption(93).
Conclusions
It is important to correlate the effects produced experimentally, which are generally based on the use of concentrations much higher than those found naturally in the feed, and generally continuously for long periods, and not to underestimate the presence of more than one mycotoxin that can enhance the individual effect of each of them. It should also be considered that, in the case of poultry, pathways of absorption, distribution and metabolism of toxins may be different from other species. As already described, fusariotoxins can lead to large economic losses in production, either due to the alteration of production parameters or their alternate effect on intestinal integrity and immune response, therefore, it is necessary to make a correct diagnosis if the presence of mycotoxins in the feed is suspected.
Conflict of interest
The authors declare no conflict of interest.
Literature cited: