https://doi.org/10.22319/rmcp.v13i2.6032
Revisión bibliográfica
Función ovárica y respuesta a la sincronización del estro en ganado Criollo en México. Revisión
Elizabeth Pérez-Ruiz a
Andrés Quezada- Casasola b
José María Carrera-Chávez b
Alan Álvarez-Holguín a
Jesús Manuel Ochoa-Rivero a
Manuel Gustavo Chávez-Ruiz a
Sergio Iván Román-Ponce a*
a INIFAP. Centro de Investigación Regional Norte-Centro. CE La Campana. Km 33.5 Carr. Chihuahua – Ojinaga. 32910. Aldama, Chihuahua. México.
b Universidad Autónoma de Ciudad Juárez. Departamento de Ciencias Veterinarias. Ciudad Juárez, Chihuahua, México.
* Autor de correspondencia: roman.sergio@inifap.gob.mx
Resumen:
Hoy en día, las biotecnologías reproductivas han hecho posible conservar y aprovechar los recursos zoogenéticos. Una de estas tecnologías son los programas de sincronización de estros, mismos que permiten programar la época para el empadre acorde a la disponibilidad de forraje o el nacimiento de los becerros por fines comerciales. Otra aplicación es la reducción del intervalo parto primera ovulación, mediante protocolos que facilitan el uso de inseminación artificial. Los bovinos criollos son un recurso genético valioso, debido a su rusticidad y adaptabilidad a condiciones ambientales difíciles; son resistentes a parásitos, aprovechan los recursos forrajeros disponibles y se reproducen en sistemas con poca o nula suplementación. En México, los primeros estudios de sincronización de bovinos criollos sugieren que las vacas criollas no responden adecuadamente a los protocolos hormonales y se obtienen porcentajes de gestación inferiores a los obtenidos en otras razas. Lo anterior, dio origen a una serie de investigaciones en fisiología reproductiva y uso de biotecnologías en ganado criollo. El objetivo de esta revisión es recabar la información existente sobre el uso de protocolos de sincronización de estros y ovulación en bovinos criollos de México; con la finalidad de poder identificar las líneas de investigación necesarias para el desarrollo de protocolos de sincronización de estros y ovulación adecuados para el ganado criollo.
Palabras clave: Bovinos carne, Recursos zoogenéticos, Inseminación artificial, Ovulación.
Recibido: 11/08/2021
Aceptado: 03/01/2022
Introducción
Los bovinos criollos son los descendientes del ganado ibérico transportado al continente americano, durante la colonización de estos países por parte de españoles y portugueses(1). Uno de los rasgos más importantes de los bovinos criollos es su adaptabilidad a condiciones ambientales difíciles; se reproducen en sistemas con fluctuaciones extremas de temperatura ambiente(2,3), son resistentes a parásitos(4) y aprovechan una variedad de plantas herbáceas, además de alternar entre pastoreo y ramoneo(1). La capacidad de adaptabilidad de una raza a ambientes variables, llamada plasticidad fenotípica, y es una cualidad de los bovinos criollos, que puede ser aprovechada en los sistemas de selección y reproducción actuales(1). Por lo cual estos bovinos son un recurso genético que podría contribuir a mejorar la productividad en un entorno desafiante(1,5). Respecto a los recursos genéticos animales, en marzo de 2018, 7,745 razas de las 8,803 razas registradas por la FAO se clasificaron como razas locales (es decir, se informó que están presentes en un solo país) y se extinguieron un total de 594 razas locales. Entre las razas locales existentes, el 26 % se clasificó como en riesgo de extinción, el 7 % no están en riesgo y el 67 % como desconocido(6). Una de las posibles causas de la disminución de la población de razas locales es porque son consideradas poco productivas, en comparación con razas especializadas (europeas y cebuinas). En general, los bovinos criollos son animales de talla pequeña y difícilmente pueden competir con otras razas de ganado especializado en la producción de carne o leche, por lo que muchos ganaderos han optado por realizar cruzamientos con estos, o sustituir a los criollos con otras razas de ganado(7,8).
Las biotecnologías reproductivas son herramientas útiles para abrir la posibilidad de conservar y aprovechar los recursos zoogenéticos(9). Con la aplicación de programas de sincronización de estros, se tiene posibilidad de elegir la mejor época para el empadre (acorde a la disponibilidad de forraje), reducir el intervalo del parto a la primera ovulación (PPO), facilitar el uso de inseminación artificial (IA) e implementar programas de mejoramiento genético(10). La manipulación del ciclo estral y la inducción de la ovulación mediante los diferentes protocolos de sincronización en conjunto con IA presentan numerosas ventajas adicionales, como lo son la producción de becerros en lotes homogéneos y la posibilidad de incrementar el precio de venta, así como facilitar el manejo nutricional y sanitario del hato(11).
Sin embargo, el uso de biotecnologías reproductivas, como la sincronización del estro y la ovulación es poco utilizada en bovinos criollos, y con resultados variables(12,13). En México, los primeros estudios relacionados con la sincronización de bovinos criollos se realizaron en la década de 1990, y en ellos se mencionó que las vacas criollas no responden adecuadamente a los protocolos hormonales(12). Por lo anterior, el objetivo de la presente revisión es recabar la información existente del uso de protocolos de sincronización de estros en bovinos criollos de México y de esta manera, poder identificar las líneas de investigación necesarias para el desarrollo de protocolos de sincronización de estros y ovulación adecuados para el ganado criollo, lo que permitirá direccionar los esfuerzos de investigación y desarrollo.
Inseminación artificial a tiempo fijo en bovinos
Con el uso de hormonas exógenas y sus análogos se busca controlar el desarrollo folicular, la regresión del cuerpo lúteo (CL) y la ovulación, para posteriormente realizar IA a estro detectado o IA a tiempo fijo (IATF). Con los protocolos hormonales para IATF se logra: 1) reducir el número y frecuencia de manejo del ganado, y 2) eliminar la necesidad de detección de estros para realizar la IA, por lo que son los más usados en ganado de carne(14).
Los tratamientos de sincronización de estros se basan principalmente en el uso de dos tipos de hormonas, progestágenos (principalmente progesterona, P4) y análogos de prostaglandina F2α (PGF2α). En los protocolos de sincronización de la ovulación, adicionalmente se utilizan análogos de estradiol (E2) y hormona liberadora de gonadotropinas (GnRH)(15). La preferencia en el uso de estos tratamientos para IATF puede estar limitado en los países en los que existen restricciones del uso de E2(16) tales como Estados Unidos de América, Australia, Reino Unido, entre otros. El porcentaje de gestación que se puede obtener con estos protocolos de IATF varía en un rango de 40 a 60 %(15). El establecimiento de la gestación depende en gran medida de la correcta función del CL, la adecuada señalización del embrión para el reconocimiento materno de la gestación y del ambiente oviductual y uterino que favorezca el desarrollo del embrión. Estos factores se ven modificados en parte por las condiciones preovulatorias, es decir, existe un efecto de la capacidad esteroidogénica del folículo, y la consiguiente competencia del ovocito(17). Otros factores críticos para el éxito de los protocolos de sincronización son el estado fisiológico de los animales (vaquillas prepúberes, vacas cíclicas o anéstricas), condición corporal (CC)(18), nutrición, calidad del semen, destreza del inseminador(11) y tiempo de inseminación posterior al tratamiento hormonal(19).
Por ejemplo, las hembras con CC baja (usualmente ≤ 4 en escala 1 a 9)(11), tienden a estar en anestro y, por lo tanto, presentan porcentajes de gestación bajos en comparación con hembras con mejor CC(18). Adicionalmente, las vaquillas de primer parto son más sensibles a la pérdida de peso y a una baja CC, en comparación con las vacas multíparas(18). La ingesta de nutrientes y el balance energético, antes y después del parto, afectan la duración del anestro postparto y el intervalo del parto a la concepción, así como el porcentaje de gestación(11). Cuando se realiza la IA, la eficiencia del inseminador se ve influenciada por la calidad y manejo del semen, así como por su habilidad técnica para depositar el semen en el lugar correcto(11). Además, el manejo correcto y evaluación de las pajillas de semen es fundamental para asegurar la calidad del material utilizado, pues influye directamente en la tasa de fertilización y, por ende, en la tasa de gestación. Previo a la IA, se recomienda hacer la evaluación del semen que será utilizado, según las características seminales más importantes(11). Por lo anterior, es imperativo que los técnicos inseminadores sean lo suficientemente capacitados para guiar la pistola de IA a través del cuello uterino, para depositar el semen completamente en la entrada del cuerpo uterino(11).
Protocolos convencionales para IATF basados en estradiol y progesterona en bovinos
El protocolo basado en el uso de P4 y E2 es conocido como convencional. La P4 simula la función del CL, inhibiendo la liberación de pulsos de GnRH/LH. El E2, aplicado al inicio del tratamiento con P4, provoca la ovulación del posible folículo dominante existente y la atresia del resto de los folículos subordinados. Con el surgimiento de una nueva oleada folicular, entre tres y cinco días después, se busca la presencia de un folículo dominante nuevo y un ovocito viable al finalizar el tratamiento con P4(20). La administración de E2 al final del tratamiento con P4 induce, de igual forma que la primera aplicación, una retroalimentación positiva sobre el hipotálamo para la liberación de GnRH y el consiguiente incremento en la frecuencia de pulsos de LH, con lo cual se sincroniza y se reduce el tiempo en que se presenta la ovulación, para realizar la IATF(20).
El protocolo convencional, consiste en la inserción de un CIDR y de la administración de BE (a dosis total de 2 mg, vía IM) el día de inicio del tratamiento con P4 (día 0)(16). El 17 β estradiol (E-17β) tiene una vida media menor que el BE, por lo que este último ha mostrado ser más eficiente en los protocolos de IATF(20). Adicionalmente, se administra PGF2α al momento del retiro del CIDR, para asegurar la regresión del CL que se haya formado después de la ovulación posterior a la primera aplicación de E2 (día 7, 8 o 9, en caso de existencia de un CL)(21). Para sincronizar la ovulación se puede aplicar una dosis total de 1 mg de BE, 24 h después de terminado el tratamiento con P4, y realizar la IATF 30 a 34 h después de aplicar la segunda dosis de BE(22). Una modificación a este protocolo consiste en administrar una dosis total de 0.5 o 1 mg de CE, al momento de retirar el dispositivo con P4, con lo cual se simplifica el manejo del ganado, con IATF 48 a 56 h después(15,23). El CE tiene una vida media mayor que el BE, por lo cual su uso permite disminuir la cantidad de manejos realizados al ganado(23). Otra opción es aplicar una dosis de 100 µg de GnRH, 54 h después del retiro del CIDR (al momento de realizar la IATF), con lo cual se induce la ovulación del nuevo folículo dominante, en caso de que no haya ovulado en forma espontánea(14). Una alternativa más a este protocolo es la administración de gonadotropina coriónica equina (eCG) al momento de terminar el tratamiento con P4(24) (Figura 1). La eCG se une a los receptores de FSH y LH, provocando el incremento de la tasa de crecimiento del folículo dominante, estimula la expresión de enzimas esteroidogénicas, principalmente de E2 folicular, con la consiguiente presentación del pico preovulatorio de LH. Adicionalmente, la eCG, produce un aumento en el diámetro del CL e incrementa la producción de P4 posterior a la IA(25). Este efecto de la eCG es más marcado en hembras con baja CC (y que ganan peso durante la época de empadre) y en anestro postparto (PP)(24).
En ganado de carne, se usan dosis de 300-400 UI de eCG, estas dosis son las más usadas tanto en protocolos convencionales(21) como en protocolos basados en GnRH(26,27). La eCG puede promover el crecimiento de uno o varios folículos en la misma oleada, por lo que no solo se logra un mayor diámetro folicular, sino también el incremento de la tasa de ovulación en vacas tratadas con esta hormona, cuando se usan dosis altas (400-600 UI)(28). La presentación de partos múltiples en ganado de carne se considera indeseable, por lo que el incremento de la tasa de ovulación por efecto de la eCG, es controversial(27). Estos efectos de la eCG son más evidentes durante el anestro PP, en vacas amamantando y con baja CC(24), debido a que la secreción de GnRH/LH se encuentra disminuida durante esta etapa(28). Sin embargo, en vacas con buena CC, no se observa un efecto benéfico(26).
Figura 1: Representación esquemática de protocolos de sincronización basados en progesterona (P4) y esteres de estradiol (BE: benzoato de estradiol; CE: cipionato de estradiol) (PGF2α: prostaglandina F2α; GnRH: hormona liberadora de gonadotropinas; eCG: gonadotropina coriónica equina)
En los tratamientos CE (Cipionato de estradiol), GnRH (Hormona liberadora de gonadotropinas) y eCG (gonadotropina coriónica equina) se sustituye la segunda dosis de BE (benzoato de estradiol) por la hormona respectiva, al momento del retiro del CIDR (dispositivo intravaginal de liberación de progesterona), para simplificar el manejo del ganado.
Protocolos para IATF basados en GnRH y PGF2α
La aplicación inicial de un análogo de GnRH provoca la liberación de LH en forma de pico preovulatorio y, por tanto, la ovulación de un posible folículo dominante presente, con la subsiguiente aparición de una nueva onda folicular aproximadamente dos días después(29). La administración de PGF2α 7 d después de la aplicación de GnRH induce la regresión del posible CL formado después de la aplicación de GnRH y una segunda dosis de GnRH provocará nuevamente la liberación del pico preovulatorio de LH, produciendo la ovulación de un nuevo folículo dominante en forma sincronizada(30).
El protocolo más usado para IATF en ganado lechero es conocido como Ovsynch (sincronización de la ovulación)(29). Este protocolo requiere del manejo del ganado en tres ocasiones para aplicar las hormonas y una cuarta vez para realizar la IATF (Figura 2), por lo que es poco práctico para su uso en ganado productor de carne(31,32). La alternativa para este tipo de ganado es el protocolo CO-Synch(33) (la segunda dosis de GnRH es aplicada al momento de la IATF), en el que se reduce el número de ocasiones en las que se maneja al ganado(32). Sin embargo, con este protocolo, 5 a 20 % de las hembras en anestro PP, presentan estro antes o inmediatamente después de la aplicación de PGF2α, por lo que se obtiene una tasa de gestación menor que con el protocolo Ovsynch(34). La inserción de un CIDR, entre la primera administración de GnRH y la aplicación de PGF2α (Figura 2), incrementa la tasa de gestación con este protocolo(34,35). El uso del CIDR previene la ovulación antes y después de la aplicación de PGF2α, causada por luteólisis espontanea del CL, y como resultado se obtienen estros más sincronizados con el momento de la IATF, por lo que con el protocolo CO-Synch + CIDR se obtienen mayores tasas de gestación(34,35). El éxito de este protocolo depende en gran medida del porcentaje de hembras que ovulan después de la primera dosis de GnRH(14). Si se logra inducir la ovulación del folículo dominante, la emergencia de la siguiente onda folicular y ovulación será sincronizada(22).
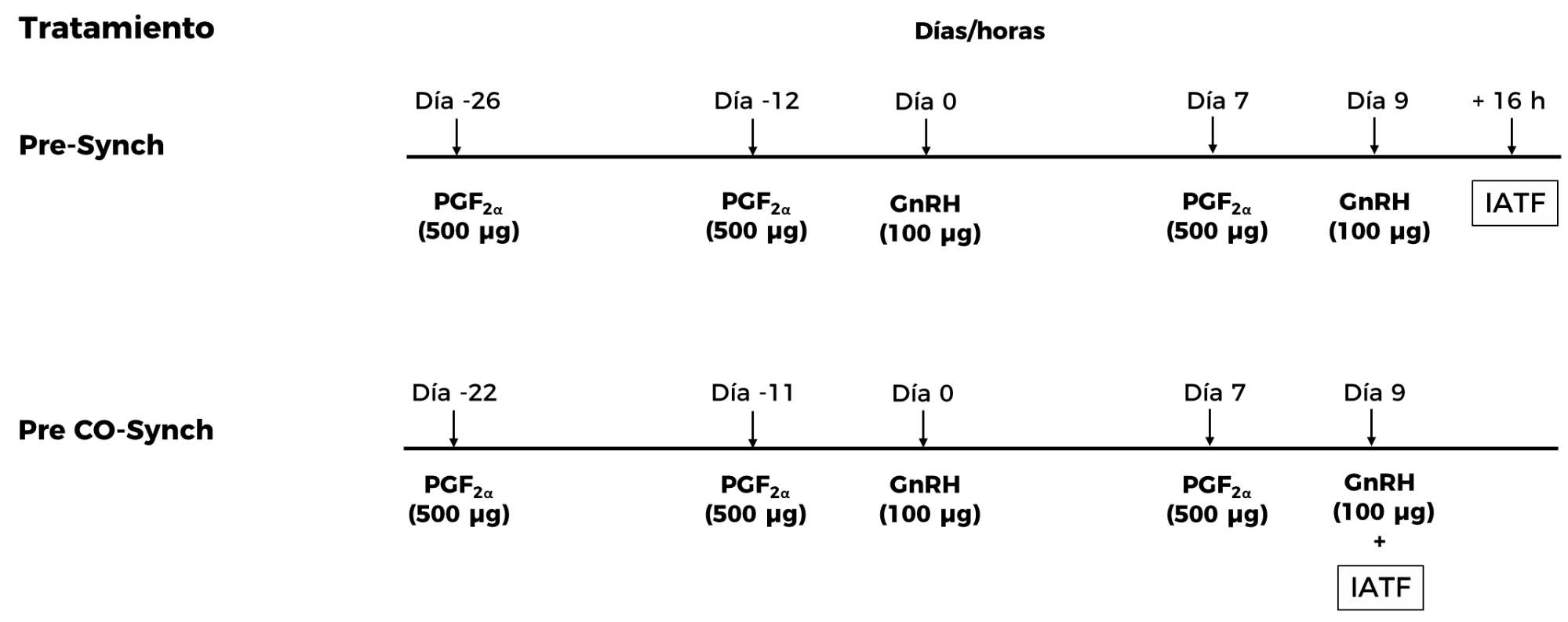
En ganado lechero, una alternativa más para aumentar el porcentaje de vacas gestantes en los protocolos Ovsynch es la pre-sincronización con una o dos dosis de PGF2α(36), con diferencia de 14 días entre cada dosis, y la aplicación de la primera dosis de GnRH, 12 a 14 días después de la segunda dosis de PGF2α, este protocolo es conocido como Pre-Synch (Figura 3). El objetivo de esta pre-sincronización es que las vacas se encuentren entre los días 5 y 12 del ciclo al momento de iniciar el tratamiento con GnRH(36,37). En ganado productor de carne, la pre-sincronización es poco práctica, dado que este protocolo implica manejar al ganado un mayor número de ocasiones y no aumenta el porcentaje de hembras gestantes después de la IATF(37). Como resultado del estrés que se produce al introducir a las vacas productoras de carne en los corrales y mangas de manejo, los animales experimentan una respuesta de lucha o huida, lo que activa el eje adrenal y la liberación de hormonas del estrés (catecolaminas y glucocorticoides), que tienen un efecto negativo sobre el eje reproductivo(38).
Figura 2: Representación esquemática de los protocolos Ovsynch, Co-Synch y CO-Synch+CIDR para IATF (GnRH: hormona liberadora de gonadotropinas; PGF2α: prostaglandina F2α; IATF: inseminación artificial a tiempo fijo)
Figura 3: Representación esquemática de los protocolos Pre-Synch usados en ganado lechero y Pre CO-Synch desarrollado para ganado productor de carne (GnRH: hormona liberadora de gonadotropinas; PGF2α: prostaglandina F2α; IATF: inseminación artificial a tiempo fijo)
Protocolo Co-Synch de 5 días
El protocolo Co-Synch de 5 días con IATF 72 h después (Figura 4), se basa en la idea de que incrementar el periodo en el cual el folículo dominante se desarrolla en presencia de gonadotropinas, puede incrementar el porcentaje de hembras gestantes después del tratamiento hormonal(39). Los principales cambios en este protocolo son: 1) la reducción del tratamiento con P4 de 7 a 5 días, para evitar los efectos adversos de los folículos persistentes sobre la fertilidad de las vacas que no ovulan con la primera dosis de análogo de GnRH (dosis total de 100 µg de gonadorelina, IM), y 2) prolongar el período desde el retiro de la P4 a la administración de GnRH, para aumentar la exposición a las concentraciones de E2 circulante antes de la ovulación(17). Con este protocolo se observa mayor diámetro del folículo dominante, incremento de la concentración de E2, así como mayor producción de P4 posterior a la ovulación, necesaria para la gestación(40).
Figura 4: Protocolos CO-Synch 5 días y J-Synch
En estos protocolos se prolonga el proestro (incremento del periodo entre el fin del tratamiento con Progesterona P4) y la IATF (PGF2α: prostaglandina F2α; GnRH: hormona liberadora de gonadotropinas; BE: benzoato de estradiol; IATF: inseminación artificial a tiempo fijo)
Protocolo J-Synch
Este protocolo se basa en el uso de P4 y BE, con una mayor duración del proestro que con el tratamiento convencional(41). El protocolo inicia con la administración de una dosis total de 2 mg de BE, en el momento de la inserción de un dispositivo con P4, que se retira 6 días después. Al momento del retiro del dispositivo con P4 se aplica una dosis única de PGF2α. De forma adicional, se aplica 100 µg de GnRH al momento de la IATF, 72 h después (día 9, Figura 4). Similar al protocolo CO-Synch 5 días, con la prolongación del proestro (95-97 h) se observa un incremento en las concentraciones de E2 (antes de la ovulación) y P4 (después de la ovulación), así como un mayor porcentaje de gestación (cuando las hembras se encuentran en buena CC), en comparación con el protocolo convencional Co-Synch(42). Estudios recientes indican que la duración del proestro es decisiva para el establecimiento de la gestación. Con un proestro más largo, se incrementa la producción de E2 antes de la ovulación. Entre las funciones del E2, se encuentran la modificación de la morfología celular, secreción y regulación de receptores esteroidales, que favorecen la implantación del conceptus en el útero(43).
En el Cuadro 1 se presentan algunos resultados obtenidos para el porcentaje de preñez, con distintos protocolos de sincronización.
Cuadro 1: Porcentaje de gestación obtenido con diferentes protocolos para IATF en bovinos
Tipo de Protocolo† | Tipo racial‡ | Gestación (%) | Referencia |
Basados en P4 y E2 | |||
Convencional (BE) | BI, BT | 30.0-40.6 | (25, 44-46) |
Convencional (CE) | BI, BT | 34.7-54.0 | (16, 42, 44, 47, 48) |
J-Synch | BI/BT, BT | 47.0-67.9 | (16, 41, 42, 47, 49) |
Convencional BE + eCG | BI | 36.8-57.5 | (25, 46, 50, 51) |
Convencional CE + eCG | BI, BT | 50.3-61.8 | (16, 28, 43, 48, 52) |
J-Synch + eCG | BI/BT, BT | 53.0-60.4 | (16, 43, 47, 53) |
Basados en GnRH y PGF2α | |||
Ovsynch | BT | 32.5-57.0 | (33, 36, 37, 54, 55) |
Pre-Synch | BT | 27.0-49.6 | (36 , 37, 55– 57) |
CO-Synch | BT | 26.7-53.3 | (33- 35, 54, 58) |
CO-Synch +CIDR | BT | 50.0-55.1 | (26, 34, 35, 58- 60) |
CO-Synch 5 d | BT | 44.4-59.7 | (61- 63) |
CO-Synch 5 d + CIDR | BT | 48.0-63.9 | (59, 60, 63- 66) |
CO-Synch +CIDR+ eCG | BT | 43.0 | (26) |
CO-Synch 5 d +CIDR+ eCG | BT | 42.9 | (67) |
†CIDR= dispositivo intravaginal de liberación de progesterona; P4=Progesterona; E2= Estradiol; PGF2α= prostaglandina F2α; GnRH= Hormona liberadora de gonadotropinas; BE= benzoato de estradiol; eCG= gonadotropina coriónica equina; CE= Cipionato de estradiol.
‡ BT: Bos taurus taurus; BI: Bos taurus indicus.
Bovino criollo en México
El ganado criollo en México desciende de los primeros ejemplares traídos por los españoles durante la época de la Colonia en el siglo XVI(68). Estos bovinos desarrollaron cualidades de adaptación al medio en zonas aisladas y de difícil acceso, lo que contribuyó a la formación de grupos raciales(69,70). En México, de las 53 razas de bovinos integradas en la base de datos sobre la biodiversidad de los animales domésticos publicada por la FAO(71), destacan como razas locales el Chinampo, de Baja California(72,73); el Coreño, de la Sierra Madre Occidental (74,75); el criollo de las Montañas del Norte, también llamado criollo de rodeo o criollo Rarámuri(70); el Mixteco(76); el criollo del Golfo(77); el criollo de la región central de Chiapas(78) y el criollo Nunkiníen, de la Península de Yucatán(79). A partir de 1965, también se introdujeron individuos de las razas criollo lechero tropical y Romosinuano, provenientes de Estados unidos y Costa Rica, respectivamente(80).
El bovino criollo Rarámuri (CR), también llamado “Corriente”, está adaptado a la región norte del país, principalmente en la Sierra Tarahumara, en el estado de Chihuahua(70). Son animales de tamaño pequeño y cornamenta grande; su principal fin zootécnico es para las actividades deportivas de rodeo, por lo que es exportado en grandes cantidades a Estados Unidos(7). La fisiología reproductiva de esta raza de bovinos criollos de México es la más investigada hasta ahora, ésta incluye la caracterización del ciclo estral, actividad ovárica, comportamiento estral, perfiles hormonales(81-83) y desarrollo, y evaluación de protocolos de sincronización e IA(12,13).
Protocolos de sincronización de estros y ovulación en ganado criollo Rarámuri
En uno de los primeros estudios en bovinos CR, se utilizó un protocolo convencional para IATF, con el uso de CIDR (con 1.9 mg de P4) por 7 días, más la aplicación de 1 mg de β-estradiol al inicio del tratamiento con P4; administración de 30 mg de PGF2α, por vía IM, al retirar el CIDR; y 24 h después de realizar el retiro del CIDR se aplicó otra dosis de 1 mg de β-estradiol. La IATF se realizó 54 h después del retiro del CIDR. Con este protocolo se observó un porcentaje de gestación de 9.09 %, a pesar de que 100 % de las vacas presentaron signos de estro. En este estudio, realizar la administración de una dosis de 50 mg de P4, por vía IM, al momento del retiro del CIDR, en conjunto con la aplicación de β-estradiol, no mejoró el porcentaje de gestación (con ambos tratamientos se obtuvo 9.09 %). El bajo porcentaje de gestación obtenido con este protocolo fue atribuido a una variación en el tiempo a la ovulación(12). Las vacas presentaron estro entre las 36 y 43 h después de la aplicación de la PGF2α. Los autores mencionan que haber dejado pasar tanto tiempo entre el inicio del estro y la IA fue un factor determinante. Cabe destacar que las vacas que presentaron estro entre 36 y 37 h después del retiro del CIDR, no resultaron gestantes, mientras que todas las vacas que presentaron estro alrededor de las 43 h después del retiro del CIDR, si quedaron gestantes. También se menciona que en vacas CR se presentan estros anovulatorios, por lo que esto contribuye a que el porcentaje de gestación sea bajo(12).
En otro estudio con vacas CR, el tiempo de inicio del estro inducido con PGF2α a la ovulación fue de 46.2 ± 8.2 h y de 37.6 ± 6.0 h en un estro natural (todas las vacas presentaron estro en un lapso de 24 a 60 h), con una proporción significativa de vacas que ovularon en el intervalo de 24 a 35 h, en el estro natural (8 vacas de un total de 22). En ambos tipos de estro, el mayor porcentaje de vacas ovularon en el intervalo de 36-47 h (12 vacas en el estro natural y 11 vacas en el estro inducido)(82). Esta diferencia en los tiempos del inicio del estro a la ovulación, en las hembras CR, respecto a los bovinos Bos taurus taurus, debe ser considerada para mejorar el porcentaje de gestación con los protocolos de IATF, o considerar la posibilidad de utilizar la regla “AM-PM”, para programar la IA, como lo mencionan Zárate-Martínez et al(12). No obstante, realizar este tipo de manejo, que requiere detección de estros, es poco práctico para los productores de bovinos CR.
El patrón de crecimiento de los folículos ováricos es en oleadas; en cada ciclo estral se pueden presentar de una a cuatro oleadas(84-86). El número de oleadas foliculares en cada ciclo es variable, siendo lo más común dos a tres oleadas(85,86). En hembras CR, hay un mayor porcentaje de vacas con dos oleadas foliculares (77.3 %)(81). Las hembras que presentan dos oleadas foliculares por ciclo ovulan en promedio 6.2 h antes que las vacas con tres oleadas(83). Además, la tasa de crecimiento folicular en la raza CR (0.6 ± 0.2 mm día-1)(82) es menor que en bovinos Bos taurus indicus (0.9 ± 0.1 mm día-1) y que en otras razas de Bos taurus taurus (1.1±0.1 mm día-1)(87). Al respecto, Quezada et al(82) mencionan que, para optimizar la respuesta a los protocolos de sincronización en vacas CR, es necesario modificar el tiempo entre el tratamiento hormonal y la IA, por lo que sugieren realizar la IA ~28 h después del inicio del estro(82).
Uso de eCG en ganado criollo
En ganado Bos taurus taurus, la aplicación de eCG en protocolo para IATF, ayuda a incrementar el ritmo de crecimiento y diámetro folicular, así como la producción de P4, en hembras con baja CC(24). Al evaluar el uso de una dosis de 400 UI de eCG al momento de la IATF (56 h después del retiro del CIDR) en vacas CR, con el uso de un protocolo CO-Synch 8 d (aplicación de una dosis 100 µg de GnRH al insertar el CIDR + 25 mg de PGF2α al retirar el CIDR, el día 8 del protocolo + 400 UI de eCG, al momento de la IATF) no se mejoró el porcentaje de gestación (31.5 vs 46.6 %) respecto al mismo protocolo sin eCG (en este tratamiento se aplicó 100 µg de GnRH al momento de la IATF). Los autores mencionan que las hembras estaban en una CC aceptable (4.5 ± 0.2; en una escala de 1 a 9) y recibieron una buena alimentación durante el manejo reproductivo, por lo cual no se observó un efecto positivo de la eCG(88). En este mismo estudio, la suplementación con concentrado, selenio (0.95 mg Se/50 kg PV) o propionato de Ca (100 g), tampoco modificó el porcentaje de gestación(88).
En otro estudio en el que se evaluó el uso de una dosis de 500 UI de eCG en vacas CR, se obtuvo un porcentaje de gestación de 60 %, del total de hembras en el tratamiento (y 75 % de gestación, respecto a las que mostraron signos de estro)(13). El protocolo hormonal que se utilizó en este estudio consistió en el uso de un CIDR por 7 días + 2.76 mg de BE al inicio de tratamiento con P4 (día 0) + 25 mg PGF2α (día 7) + 1 mg de CE o 500 UI de eCG 24 h después de retirar el CIDR (día 9); la IA se realizó 12 h después del inicio del estro. El porcentaje de gestación en el grupo de vacas tratadas con CE fue de 27.3 %. Cabe destacar que, en este estudio, 100% de las vacas tratadas con CE y 80% de las tratadas con eCG mostraron signos de estro en respuesta al protocolo de IA y todas las hembras en ambos tratamientos ovularon. La respuesta de las vacas a ambos tratamientos fue similar para las variables: respuesta a estro, porcentaje de ovulación y diámetro folicular máximo(13). Al evaluar estos mismos protocolos en novillas CR, la respuesta a estro fue menor en ambos tratamientos, 89.5 % y 25.0 % para los tratamientos CE y eCG, respectivamente. El porcentaje de novillas que presentaron estros silenciosos fue de 10.5 % para las novillas del grupo tratado con CE y 75.0 % de las tratadas con eCG. La presentación de estros silenciosos fue atribuida a un menor crecimiento folicular que el observado con el uso de CE. Debido a que solo las hembras que mostraron signos de estro fueron inseminadas, el porcentaje de gestación con el uso de eCG fue de solo 10 %, respecto al total en el grupo (40 % de las que presentaron estro y fueron inseminadas)(13). No obstante, 100 % de las vaquillas tratadas con eCG ovularon. En este estudio, tanto en vacas como en vaquillas, el estro se presentó en menor tiempo con CE (vacas 24.9 ± 2.8 h y vaquillas 25.8 ± 2.9 h) en comparación con el tratamiento de eCG (31.5 ± 2.8 y 30.6 ± 2.9). Adicionalmente, los autores mencionan la posibilidad de usar IATF exitosamente, en vacas multíparas con el uso de eCG, puesto que, con este protocolo, el inicio del estro se agrupó entre las 24 y 36 h después de retirar el CIDR (31.5 ± 2.8 h en promedio) y el mayor porcentaje de vacas fueron inseminadas 36 a 48 h después de terminar el tratamiento con P4. Además, los resultados de respuesta al estro y porcentaje de gestación obtenidos en este estudio son superiores a los obtenidos por Zárate et al(12) y Sánchez-Arciniega et al(88) en esta misma raza de bovinos.
Reinicio de la actividad ovulatoria/cíclica postparto
El anestro postparto (PP) se caracteriza por ausencia de ovulaciones después del parto. En este periodo los folículos ováricos inician el crecimiento, pero ninguno es capaz de ovular, al menos durante las primeras semanas(89). Esto es debido en parte a la ausencia de LH, y frecuentemente, la primera ovulación no es precedida por manifestación de signos de estro; el CL puede tener una vida media reducida, menor tamaño y actividad esteroidogénica limitada(90). Para disminuir el efecto negativo del amamantamiento se ha propuesto realizar destete precoz (DP, pocos días después del parto); amamantamiento controlado o restringido (AR, consiste en permitir el amamantamiento en periodos cortos en el día); o destete temporal (DT, separar al becerro de la madre durante algunos días)(91). La técnica de AR incrementa la proporción de vacas que manifiestan signos de estro durante los primeros 100 días postparto y reduce el intervalo parto primera ovulación, sin afectar el crecimiento de los becerros(91).
En ganado CR, con la estrategia de AR (inicio al día 76 PP), previo al protocolo hormonal para IATF, 81.4 % de las vacas ovularon dentro de un periodo de observación de 22 días después de iniciado el AR, pero solo 11.1 % de ellas manifestaron signos de estro, es decir presentaron ovulaciones silenciosas. En este estudio, la manifestación de la conducta estral solo se observó en las hembras con mejor CC (4-5, regular a buena), mientras que las vacas con CC mala (2-3, en una escala de 1-9), no ovularon antes del tratamiento de sincronización(12).
En otro estudio en ganado CR, al evaluar la pérdida de peso durante la lactancia, las vacas que fueron tratadas con un esquema de DP a partir de los 68 ± 3.8 d PP, perdieron menos peso que aquellas que permanecieron con las crías (destete normal a los 180 ± 10.2 días) durante el periodo de evaluación. En las hembras con DP, la pérdida de peso fue de 4.8 kg, mientras que las hembras del grupo con destete normal perdieron 18.9 kg durante el periodo de evaluación (68-180 días PP)(92). Como se ha mencionado, al igual que en otras razas bovinas especializadas(11), la CC en el ganado CR es una limitante para reestablecer la actividad reproductiva después del parto, y para el establecimiento de la gestación(92). El ganado CR en pocas ocasiones es suplementado, por lo que no se tiene control sobre su condición corporal(88). Por lo anterior, realizar estrategias como AR o DP en ganado criollo, en conjunto con la implementación de protocolos para IATF podría ser de utilidad para mejorar el porcentaje de gestación.
Sincronización de estros en otras razas de ganado criollo en México
La información del desempeño reproductivo en respuesta a protocolos de sincronización de estros e IA en otras razas de bovinos criollos en México es limitado. En vacas criollo Coreño de Nayarit, con la sincronización con un implante de Norgestomet por 9 días + 280 UI de eCG, se obtuvo 80 % de respuesta a estro y un porcentaje de gestación de 60 %(93).
En la raza Chinampo, se evaluó el uso de dos dosis de PGF2α (con 11 días de diferencia entre cada aplicación) para inducir el estro y evaluar el comportamiento estral en presencia del semental(94). En presencia del toro, se encontró mayor interacción entre las 30 y 60 h después de la segunda dosis de PGF2α. Las vacas expuestas al semental iniciaron estro en menor tiempo y también tuvieron una duración del estro menor que aquellas que permanecieron aisladas del toro (10.7 ± 1.1 h vs 16.3 ± 2.6 h).
En vaquillas puras y mestizas de la raza criollo lechero tropical, se encontró una respuesta de estro de 94.1 % y porcentaje de gestación de 68.8 %; con la adición de una dosis de 500 UI de eCG, al d 10, de un protocolo con implante subcutáneo (con 3 mg de norgestomet, por 12 días + 5 mg de valerato de estradiol, VE, por vía IM el día 0). Mientras que con una dosis de 0.25 mg de GnRH, 24 h después del retiro del implante (implante con 3 mg de norgestomet, por 12 días + 5 mg de VE, por vía IM el día 0), se obtuvo 76.4 % de hembras que mostraron estro y 46.2 % de ellas quedaron gestantes(95). En este protocolo, se realizó la aplicación de una dosis de 15 mg de PGF2α, vía IM, 10 días antes de colocar el implante subcutáneo; para homogeneizar el ciclo estral.
En el Cuadro 2 se resumen los resultados obtenidos en bovinos criollos, con el uso de diversos protocolos de sincronización de estros e IA.
Cuadro 2: Resumen de porcentajes de gestación obtenidos con protocolos hormonales en bovinos criollos en México
Raza† | Protocolo‡ | Gestación (%) | Referencia | |
Vacas | CR | CIDR por 7 días (d) + 1 mg E2 + 50 mg de P4 (d 0) + 30 mg de PGF2α (d 7) + 1 mg de E2 (d 8) | 9.1 | (12) |
Vacas | CR | CIDR por 7 d-1 + 1 mg E2 (d 0) + 30 mg de PGF2α, (d 7) + 1 mg de E2 (d 8) | 9.1 | (12) |
Vacas | CR | CIDR por 7 d-1, 100 µg de GnRH + 25 mg de PGF2α (d 8) + 400 UI 56 h después de retirar el CIDR | 31.5 | (88) |
Vacas | CR | CIDR por 7 d-1, 100 µg de GnRH + 25 mg de PGF2α (d 8) + 100 µg 56 h después de retirar el CIDR | 46.6 | (88) |
Vacas | CR | CIDR por 7 d-1 + 2.76 mg de BE (d 0) + 25 mg PGF2α (d 7) + 1 mg de CE (d 9) | 27.3 | (13) |
Vacas | CR | CIDR por 7 d-1 + 2.76 mg de BE (d 0) + 25 mg PGF2α (d 7) + 500 UI de eCG (d 9) | 60.0 | (13) |
Vacas | Coreño | Implante de Norgestomet por 9 días + 280 UI de eCG | 60.0 | (93) |
Vaquillas | CR | CIDR por 7 d-1 + 2.76 mg de BE (d 0) + 25 mg PGF2α (d 7) + 1 mg de CE (d 9) | 27.3 | (13) |
Vaquillas | CR | CIDR por 7 d-1 + 2.76 mg de BE (d 0) + 25 mg PGF2α (día 7) + 500 UI de eCG (d 9) | 60.0 | (13) |
Vaquillas | CLT | 15 mg de PGF2α (d -10) + Implante subcutáneo (3 mg de norgestomet) por 12 d-1 + 5 mg VE, 500 UI de eCG (d 10) | 68.8 | (95) |
Vaquillas | CLT | 15 mg de PGF2α (d -10) + Implante subcutáneo (3 mg de norgestomet) por 12 d-1 + 5 mg VE (d 0) + 0.25 mg GnRH (d 10) | 46.2 | (95) |
†CR= Criollo Ráramuri. CLT= Criollo Lechero Tropical.‡CIDR= dispositivo intravaginal de liberación de progesterona; E2= Estradiol; VE= valerato de estradiol; PGF2α= prostaglandina F2α; GnRH= hormona liberadora de gonadotropinas; UI= unidades internacionales; BE= benzoato de estradiol; eCG= gonadotropina coriónica equina; CE= cipionato de estradiol.
Factores asociados a la respuesta reproductiva en ganado criollo
Existen diferencias fisiológicas entre los bovinos criollos y el ganado europeo especializado en producción de carne, así como con el ganado cebú. Entre estas, destacan las inherentes al funcionamiento ovárico: número de oleadas foliculares, tasa de crecimiento folicular, diámetro del folículo ovulatorio y del CL; y concentración de hormonas reproductivas y periodos entre eventos del ciclo estral (duración de fase lútea vs fase folicular; tiempo a la ovulación, duración del estro)(82,83,85). Adicionalmente, los factores externos como estado nutricional, el manejo, las relaciones sociales y jerárquicas(38), influyen en la respuesta de los bovinos criollos a los protocolos hormonales para IATF.
Comportamiento reproductivo, función ovárica y endocrinología en la hembra criolla
El ciclo estral en vacas CR tiene una duración promedio de 21.1 ± 1.2 días (rango de 19-23 días), con una fase folicular de 6-9 días y fase lútea de 12-16 días(88). El crecimiento folicular en bovinos se da en un patrón de oleadas foliculares, y la cantidad de oleadas foliculares en cada ciclo estral es variable, pero puede ir de dos a cuatro oleadas. En vacas CR, se presenta un mayor porcentaje de hembras con dos oleadas foliculares (77.3 %), y en menor porcentaje hembras con tres ondas foliculares por ciclo (22.7 %)(82). En hembras Bos taurus taurus(84), Bos taurus indicus(85) hembras criollas tailandesas(86), también se ha observado un mayor porcentaje de hembras con dos oleadas foliculares por ciclo. Contrario a estos hallazgos, en ganado criollo Caqueteño de Colombia(9), en vaquillas criollas tendencia lechera en Ecuador(96), y criolla de la zona altoandina de Perú(86), existe un mayor porcentaje de hembras con tres oleadas foliculares (Cuadro 3). Sin embargo, hasta el momento no se cuenta con información publicada de la dinámica folicular en otras razas de bovinos criollos de México.
Cuadro 3: Número de oleadas foliculares en el ciclo estral en bovinos criollos, Bos taurus taurus y Bos taurus indicus
Raza | Numero de ondas foliculares (%) | ||
2 | 3 | 4 | |
Rarámuri de Chihuahua, México (82) | 77.3 | 22.7 | |
Caqueteño de Colombia (9) | 33.3 | 66.6 | |
Criolla zona altoandina del Perú (86) | 16.0 | 78.0 | 6.0 |
Criolla altiplano de Ecuador (96) | 44.4 | 55.6 | |
Tailandés nativa (97) | 70.0 | 30.0 | |
Nelore (85) | 83.3 | 16.6 | |
Holstein (84) | 81.0 | 19.0 | |
La caracterización del ciclo estral y dinámica folicular en diferentes razas de bovinos ha permitido observar diferencias y similitudes entre ellas (Cuadro 4). En el ganado criollo Caqueteño y Nelore, las hembras con tres oleadas foliculares tienen ciclos estrales más largos que las hembras con dos oleadas(9,85). En las vacas CR, la duración del ciclo estral, fase folicular y fase lútea, es similar entre hembras con dos y tres oleadas foliculares, pero las hembras con dos oleadas foliculares, ovulan 6.2 h antes que aquellas con tres oleadas foliculares(82,83). Los folículos ovulatorios de hembras CR con dos oleadas, crecen a una tasa menor de aquellas con tres oleadas (0.5 ± 0.04 vs 0.9 ± 0.08, respectivamente), mientras que el diámetro máximo del folículo ovulatorio es similar entre ambos patrones de crecimiento (10.5 ± 0.2 vs 10.0 ± 0.4, respectivamente)(83). El diámetro máximo del CL en vacas CR con dos y tres oleadas foliculares es similar (13.0 ± 1.0 vs 13.2 ± 1.7 mm, respectivamente)(83). El tamaño del CL y la producción de P4 en vacas CR también es menor a la de otras razas de ganado, y es posible que estas diferencias sean adaptaciones a las condiciones ambientales y nutricionales que los bovinos criollos han desarrollado para sobrevivir en ambientes difíciles(83).
Cuadro 4. Dinámica folicular y ovulatoria en bovinos con dos y tres oleadas foliculares.
Raza | ||||||||||
Rarámuri de México (83) | Criolla de Ecuador (96) | Caqueteño de Colombia (9) | Nativa tailandesa(97) | Holstein (84) | ||||||
Numero de ondas | 2 | 3 | 2 | 3 | 2 | 3 | 2 | 3 | 1 | 2 |
Duración ciclo estral, días | 21.1±0.3 | 21.4±0.6 | 20.3±0.0 | 23.6±0.0 | 20±0.6 | 22±0.5 | 18.60±0.1 | 20.38±0.1 | ||
Primer onda (no ovulatoria) | ||||||||||
Emergencia, días † | −0.5±0.2 | −0.4±0.2 | 1 | 1 | 3 | 4 | 1.53±0.1 | 1.54±0.1 | -2.0+0-1 | -0.5±0.3 |
Diámetro máximo del FD‡, mm | 8.0±0.1 | 7.6±0.3 | 13.2±2.2 | 12.2±1.7 | 8.4±1.3 | 11.7±3.3 | 7.63±0.1 | 7.67±0.1 | 17.1±0.5 | 16.0±0.4 |
Tasa crecimiento, mm día-1 | 0.5±0.03 | 0.7±0.0 | 1.0±0.1 | 1.2±0.0 | - | - | 0.65±0.0 | 0.75±0.1 | - | - |
Regresión, días | 11.0±0.8 | 7.8±0.4 | - | - | 11 | 10.0±0.5 | 8.34±0.1 | 8.79±0.1 | 13.0+0.4 | 12.2±0.5 |
Segunda onda (no ovulatoria) | ||||||||||
Emergencia, días | - | 9.6±0.5 | - | 6.6±0.0 | - | 10.0 | - | 8.38±0.1 | 9.0 | |
Diámetro máximo del FD, mm | - | 7.2±0.2 | - | 10.2±0.0 | - | 12.2±4.4 | - | 6.79±0.2 | 12.9±0.7 | |
Tasa de crecimiento, mm día-1 | - | 0.4±0.08 | - | 1.1±0.1 | - | - | - | 0.75±0.1 | - | - |
Regresión, días | - | 14.8±0.8 | - | - | 17.0±0.5 | - | 12.58±0.1 | 19 | ||
Onda ovulatoria | ||||||||||
Emergencia, días | 11.2±0.8 | 14.8±0.8 | 7.8±1.6 | 13,2±1.3 | 11.0 | 17.0 | 11.02±0.1 | 11.33±0.1 | 9.6±0.2 | 16.0±1.1 |
Diámetro máximo del FD, mm | 10.5±0.2 | 10.0±0.4 | 15.3±0.0 | 13.8±1.4 | 7.5±1.1 | 13.8±3.6 | 8.81±0.2 | 8.14±0.2 | 16.5±0.4 | 13.9±0.4 |
Tasa crecimiento, mm día-1 | 0.5±0.04 | 0.9±0.08 | 0.9±0.1 | 1.1±0.2 | - | - | 1.07±0.0 | 1.48±0.1 | - | - |
Ovulación, días | 22.0 | 22.5 | 20.0 | 23.0 | 20.0±0.6 | 22.0±0.5 | 19.44±0.1 | 21.13±0.2 | 20.4+0.3 | 22.8+0.6 |
Cuerpo lúteo (CL) | ||||||||||
Diámetro máximo del CL, mm | 13.0±1.0 | 13.2±1.7 | 21.7±1.4 | 23.5±0.6 | 11.3±4.3 | 11.2±3.2 | 13.55±0.1 | 15.14±0.1 | - | - |
Concentración máxima de P4, ng/ml | 6.5±0.1 | 6.5±0.2 | 20.6±5.4 | 20.6±3.1 | - | - | 4.13±0.1 | 4.25±0.1 | - | - |
Regresión, días | 16.3±1.6 | 16.8±1.1 | 18.0 | 20.0 | 17.0±1 | 19±0.96 | 17.11±0.1 | 19.29±0.1 | 16.5±0.4 | 19.2±0.5 |
† El momento de la ovulación (día 0), se utilizó como referente para determinar el inicio del ciclo, por lo que es posible observar la emergencia de la siguiente oleada folicular antes de la ovulación. En el Rarámuri, la evaluación de ambos ovarios se realizó cada 8 h después de iniciado el estro (día 0= día de la ovulación).
‡FD: Folículo dominante.
Interacción social y jerárquica del CR
En ganado CR, se ha observado la existencia de relaciones de dominancia entre hembras de mayor rango sobre las de menor rango jerárquico(12). La dominancia social en este tipo de ganado no solo está determinada por la edad, sino también por el tipo de cornamenta (como se mencionó, este tipo de ganado posee cuernos de gran tamaño con relación al tamaño del cuerpo, por lo cual son usados para rodeo). Hembras con defectos en los cuernos tienen dificultad para defenderse y defender a sus crías durante el amamantamiento, por lo que terminan huyendo o cediendo el espacio(12).
Otro factor que puede influir en el porcentaje de gestación es el temperamento de los animales; animales con temperamento agresivo tienden a tener un desempeño reproductivo menor al de los animales de temperamento dócil(98). El temperamento agresivo interrumpe los eventos fisiológicos necesarios para la reproducción, se asocia con una mayor síntesis y concentraciones circulantes de adenocorticotropina (ACTH) y cortisol, que pueden alterar los eventos fisiológicos clave necesarios para alcanzar la pubertad y la liberación de la oleada preovulatoria de GnRH/ LH(99). Por el contrario, un temperamento más tranquilo conduce a tasas más altas de presentación de estros y gestación, así como menos pérdidas embrionarias(100). Adicionalmente, el sistema social del ganado es muy jerárquico. La jerarquía influye la ingesta de alimento y comportamiento social(101). Además, las vacas de bajo rango adoptan una actitud pasiva como estrategia de comportamiento para reducir el estrés. La jerarquía también influye en la reproducción: las madres estresadas y de bajo rango social producen menos LH, lo que interfiere con la ovulación y el comportamiento estral(38).
Conclusiones
El uso de biotecnologías reproductivas como la sincronización de estros e IA es limitado en bovinos criollos y los resultados obtenidos son variables. Entre los factores que modifican la respuesta a la sincronización se encuentran la fisiología reproductiva propia del bovino criollo, la condición corporal y las relaciones jerárquicas. La revisión de la información existente sobre la respuesta en ganado criollo del uso de protocolos de sincronización permite proponer las líneas de investigación a desarrollar, y deben estar enfocadas en el estudio comparativo de la biología reproductiva, adecuación de protocolos hormonales tomando en cuenta las particularidades del ganado criollo y la interacción nutrición, manejo y reproducción. Finalmente, las biotecnologías reproductivas adaptadas al ganado criollo podrán ser incorporadas a los programas de conservación y aprovechamiento racional de este recurso zoogenético.
Literatura citada: