https://doi.org/10.22319/rmcp.v14i2.6003
Artículo
Composición química del rastrojo de tres cultivares de maíz esterilizados y colonizados por micelio de Ganoderma lucidum
Liz Sarahy Pérez-Martell a
Juan de Dios Guerrero-Rodríguez a*
Daniel Claudio Martínez-Carrera a
Javier Francisco Enríquez-Quiroz b
Efraín Pérez-Ramírez a
Benito Ramírez-Valverde a
a Colegio de Postgraduados-Campus Puebla. Boulevard Forjadores de Puebla No. 205, Santiago Momoxpan, 72760, San Pedro Cholula, Puebla, México.
b Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). Campo
Experimental La Posta, Km. 22.5 Carretera Federal Veracruz-Córdoba. 94277, Medellín,
Veracruz, México.
*Autor de correspondencia: rjuan@colpos.mx
Resumen:
Se evaluó la calidad nutritiva del rastrojo de dos cultivares criollos de maíz y un híbrido, colonizados por micelio de Ganoderma lucidum. El diseño experimental fue completamente al azar con arreglo factorial 3x4 con un tratamiento adicional y cuatro repeticiones. Cada cultivar tuvo rastrojo colonizado por el hongo hasta los 15 días, rastrojo en su estado natural (sin tratar), a tiempo cero después de la esterilización, a 15 días después de la esterilización y el micelio puro (adicional). Se determinó digestibilidad in vitro (DIVMS), fibra detergente neutro (FDN) y ácido (FDA), lignina y proteína cruda (PC). Los cultivares difirieron (P<0.0001) en digestibilidad, el criollo A presentó valores mayores. Los rastrojos colonizados tuvieron menor (P<0.05) digestibilidad; los rastrojos sin tratar tuvieron valores medios y los esterilizados fueron los más digestibles. La concentración de FDN, FDA, lignina, y PC difirió (P<0.0001) en los cultivares y las condiciones del rastrojo (P<0.0001). El criollo A tuvo menos FDN que los otros cultivares. Los rastrojos en su forma natural tuvieron menos FDN que los esterilizados y los colonizados. En la FDA los rastrojos en su forma natural tuvieron concentración baja, los esterilizados una concentración media y los colonizados la concentración mayor, situación que fue similar para lignina. En PC los cultivares fueron diferentes (P<0.0001), siendo los rastrojos colonizados los que tuvieron valores mayores (P<0.05). En conclusión, la colonización del rastrojo por el micelio de Ganoderma lucidum no aumentó la digestibilidad a los 15 días de colonización, lo que mejoró ligeramente fue la concentración de proteína cruda.
Palabras clave: Digestibilidad, Hongo pudrición blanca, Maíces criollos, Maíces híbridos.
Recibido: 07/06/2021
Aceptado: 07/11/2022
Introducción
El rastrojo de maíz es un ingrediente común de la dieta de los rumiantes en las zonas áridas y tropicales, principalmente cuando escasea el forraje verde durante las temporadas secas o frías que restringen el crecimiento de las plantas. Su aprovechamiento por el rumiante es limitado porque tiene concentraciones bajas de nutrientes y baja digestibilidad(1). Varios métodos de mejora de digestibilidad se han utilizado, y uno que tiene potencial es el uso de hongos comestibles o funcionales los cuales pueden transformar la estructura celular de los tejidos vegetales(2).
Los hongos de la “pudrición blanca” se han manejado como alternativa para mejorar la calidad nutricional de los rastrojos(2). Entre ellos están las especies del género Pleurotus, que tienen la capacidad de degradar la lignina mediante su sistema multienzimático que actúa sobre moléculas complejas de difícil degradación para los rumiantes(3,4,5). Algunas especies como Ceriporiopsis subvermispora, Lentinula edodes, Pleurotus eryngii o Pleurotus ostreatus, elevan el contenido de proteína cruda de los sustratos, pero provocaron una pérdida de nutrientes(6). Cepas de las especies Pleurotus florida, P. ostreatus, P. pulmonarius y P. sajor caju, se han utilizado en el tratamiento del rastrojo de maíz(7), en donde P. sajour caju incrementó mayormente la proteína cruda, la energía metabolizable y ocasionó una reducción en la concentración de la lignina.
Entre otros hongos de la pudrición blanca que tienen la propiedad de degradar lignina está Ganoderma lucidum, del cual se ha observado que comienza a degradar este compuesto entre tres a cinco meses después de la inoculación(8). No obstante, este hongo produce varios polisacáridos como la manosa, xilosa, arabinosa, galactosa, glucosa y ramnosa(9,10,11,12), además de quitina(13), que pueden incrementar el contenido de fibras. Adicionalmente, el proceso de esterilización del rastrojo con autoclave llega a ser negativo, pues se solubilizan compuestos nitrogenados del rastrojo de maíz(6). Por tanto, la cepa de hongo que se utilice es de gran importancia porque debe contar con una fuente de carbohidratos y tiempo para colonizarlo y con ello asegurar mejora de la calidad del rastrojo, además de que el consumo de este debe de ser seguro a largo plazo para los animales(14). No obstante, se desconoce el efecto que puede tener G. lucidum sobre el rastrojo de maíz en tiempos de colonización cortos, por lo que el objetivo de la presente investigación fue conocer la composición química de los rastrojos de maíz provenientes de dos cultivares criollos y un híbrido, en su estado natural, esterilizados y sujetos a diferentes tiempos de secado e inoculados con G. lucidum.
Material y métodos
Cultivares usados
Dos cultivares de maíz criollo (a las que se les denominó A y C) y el cultivar híbrido Aspros 1503® (al que se le asignó la letra B), todos de grano blanco, fueron sembrados en condiciones de temporal, en el municipio de Cuautinchán, estado de Puebla. Los cultivares se establecieron bajo un diseño en bloques al azar, con cuatro repeticiones, en una superficie de 200 x 50 m. El origen de los criollos fue de la selección de semilla de la cosecha del ciclo anterior que realizan dos productores en la misma cabecera municipal, reconocidos por conservar y mejorar el maíz de acuerdo a su conocimiento tradicional. El híbrido fue adquirido con un distribuidor de semillas Aspros en el mismo municipio, también de producción del año anterior.
Acondicionamiento del rastrojo
Al momento de la cosecha de grano que ocurrió a los 170 días después de la siembra (dds), cinco plantas de cada cultivar fueron tomadas de cada repetición. Las muestras se picaron en un molino con criba de dos centímetros. Este material constituyó la base para los tratamientos evaluados: rastrojo sin alteración, rastrojo esterilizado y secado inmediatamente, rastrojo esterilizado y mantenido húmedo durante 15 días, rastrojo esterilizado e inoculado con G. lucidum.
En cada una de las cuatro repeticiones establecidas por cultivar se obtuvieron en promedio 300 g de materia seca (MS) de rastrojo, cantidad que se dividió en cuatro partes. Una de estas partes conformó el tratamiento sin alteración alguna y las otras tres partes se sometieron a esterilización con autoclave All American® a 121.5 °C durante 25 min. Una parte del rastrojo esterilizado se sometió a un secado inmediato a 60 °C hasta alcanzar peso constante en una estufa de aire forzado Marca Thermo Scientific® Model 3478, para evaluar el efecto de lixiviación de nutrientes. La segunda parte de rastrojo esterilizado se guardó en cajas Petri (20 g MS por caja) por 15 días, para evaluar el efecto del tiempo que duró humedecido el rastrojo, en el cual pueden existir reacciones de hidrólisis que modifiquen la calidad nutritiva. De la tercera parte del rastrojo esterilizado se obtuvieron 20 g MS que se colocó en caja Petri en donde se inoculó con G. lucidum.
Preparación de la cepa del hongo e inoculación del rastrojo
La cepa de G. lucidum CP-145, se obtuvo del Centro de Biotecnología de Hongos Comestibles, Funcionales y Medicinales del Colegio de Postgraduados, Campus Puebla. Esta cepa estuvo cultivada en cajas de Petri en medio de cultivo Papa-Dextrosa-Agar (PDA) durante ocho días, tiempo suficiente de crecimiento para la obtención del micelio. Posteriormente, el rastrojo se inoculó con cinco fragmentos circulares de cinco milímetros de diámetro con micelio. El rastrojo inoculado se supervisó cada tres días hasta el día 15, que fue el tiempo en que se alcanzó el máximo de colonización micelial y es un tiempo medio en el cual algunos autores(15) han encontrado mayor actividad enzimática fúngica.
Procesamiento de los rastrojos evaluados
Todas las muestras se secaron estufa de aire forzado a 60 °C hasta alcanzar peso constante. Posteriormente, cada repetición de cada condición fue molida en un molino ciclónico FOSS TECATOR® con malla de 1 mm. Las muestras molidas se guardaron en bolsas plásticas con cierre para su conservación y posterior análisis en laboratorio para determinar la calidad nutritiva.
Dado que la presencia del hongo en el rastrojo puede modificar los contenidos de compuestos que determinan la calidad nutritiva, micelio de G. lucidum se obtuvo para conocer sus características mediante las variables de composición química que se le realizaron al rastrojo. Para esto, se utilizó medio Potato Dextrose Broth (DifcoTM) a razón de 24 g L-1, se esterilizó el medio en autoclave All American® a 121.5 °C 15 lb plg-2 por 25 min. El medio se inoculó depositando cinco círculos de 5 mm de diámetro de la colonia de G. lucidum de ocho días de edad previamente desarrollada en medio PDA. Después se incubó en agitación orbital Thermo Scientific® a 120 rpm, 27 ºC por 20 días. Posteriormente se cosechó el micelio, recuperándolo mediante un sistema de filtración con papel filtro Whatman #1 colocado en un embudo Buchner en matraz Kitazato conectado a bomba de vacío. Se obtuvo la biomasa y se secó a 60 °C en un horno de secado Felisa® en charolas de aluminio por 48 horas. Se molió en mortero de cerámica y se guardó en bolsa plástica con cierre para su posterior análisis de calidad nutritiva.
Variables evaluadas
Se determinó la fibra insoluble en detergente neutro(16) (FDN), la fibra insoluble en detergente ácido(16) (FDA), la lignina(16), la digestibilidad enzimática in vitro(17,18) y la proteína cruda (PC)(19).
Diseño experimental y análisis estadísticos
Para evaluar el comportamiento de los rastrojos de los cultivares de maíz, de los que se tuvieron tres niveles, y el acondicionamiento del rastrojo de los mismos con cuatro niveles se empleó un diseño experimental completamente al azar con arreglo factorial más un tratamiento adicional que fue el hongo puro)(20). Se verificaron los supuestos normalidad de datos y realizó el análisis de varianza respectivo. Para detectar las diferencias entre medias se utilizó la prueba de Tukey aceptando una α=0.05. El análisis de datos se realizó con el programa estadístico SAS versión 9.0(21).
Resultados
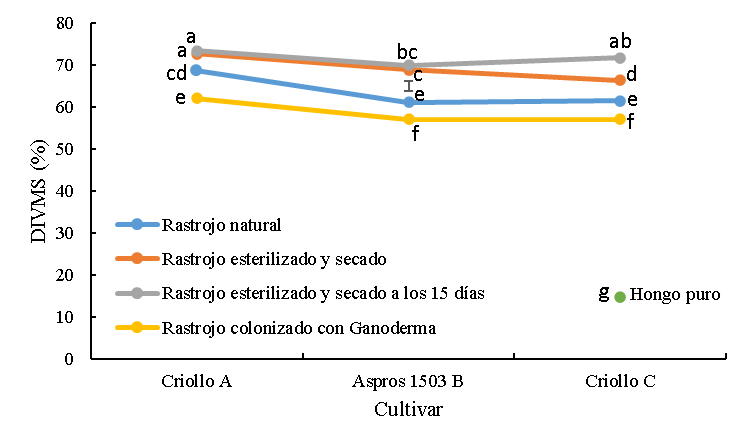
La digestibilidad in vitro de la materia seca (DIVMS) del rastrojo de los cultivares fue diferente (P<0.0001) y fue modificada mayormente por la condición a la que estuvieron sometidos los rastrojos (P<0.0001); así mismo, en dos de los cultivares se observó interacción (P<0.0001) con la condición a la que estuvieron sujetas (Figura 1).
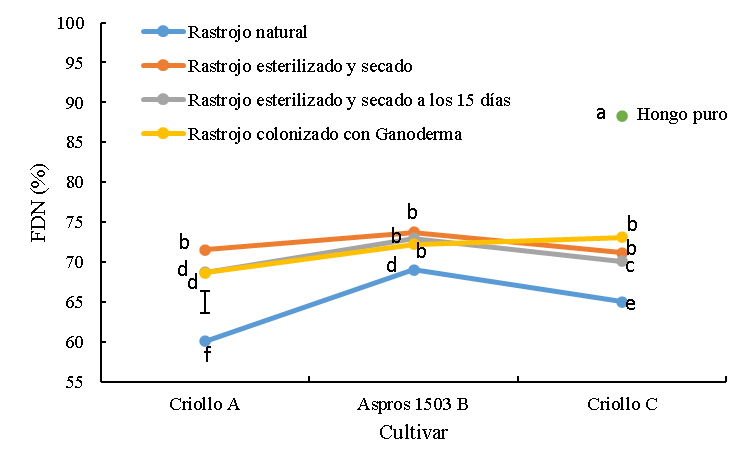
Figura 1: Digestibilidad in vitro de la materia seca (DIVMS) de rastrojos de tres cultivares de maíz sin colonizar y colonizado con Ganoderma lucidum
La barra es la diferencia honestamente significativa para la comparación entre medias.
abc Cifras con las mismas letras, son iguales (P≤0.05).
Los rastrojos sin tratar tuvieron valores intermedios de DIVMS (63.7 %) comparados con los rastrojos esterilizados y colonizados que tuvieron en promedio 70.4 % y 58.7 %, respectivamente. La variedad A fue más digestible (P<0.05) que el cultivar B Aspros 1503® y la variedad C en 7.5 y 7.1 unidades porcentuales. Los tratamientos de rastrojo que se esterilizaron y secaron inmediatamente y los esterilizados y mantenidos humedecidos por 15 días tuvieron mayor (P<0.05) digestibilidad promedio que los rastrojos sin tratar en 6.7 unidades porcentuales. En este grupo de tratamientos, la variedad A tuvo mayor (P<0.05) digestibilidad (73 %) que el cultivar B (69.3 %) y la variedad C (68.9 %).
El rastrojo colonizado tuvo valores menores (P<0.05) de digestibilidad que los rastrojos sin tratar y esterilizados, alcanzando valores de 62.0, 57.1 y 57.0 % para la variedad A, B Aspros® y variedad C, respectivamente. En este tratamiento colonizado, el criollo A mostró mayor (P<0.05) digestibilidad que los otros dos cultivares. El hongo en su estado puro tuvo la digestibilidad más baja (14.7 %) (Figura 1).
La concentración de FDN fue diferente entre cultivares (P<0.0001) y fue modificada mayormente por la condición a que estuvieron sometidos los rastrojos (P<0.0001) (Figura 2), existiendo un efecto de interacción (P<0.0001), la cual fue visible en la variedad C. En el rastrojo sin esterilizar, los cultivares fueron diferentes (P<0.05). La variedad A tuvo menor cantidad (P<0.05) de FDN respecto al cultivar B Aspros® y la variedad C, quienes tuvieron 9 y 5 unidades porcentuales más, respectivamente y resultando más alto (P<0.05) en concentración el cultivar B Aspros 1503®. Se presentó un aumento de la concentración de FDN al someter los rastrojos a la esterilización y a la colonización con el hongo respecto a los rastrojos en su forma natural (P<0.05), efecto más notable en los cultivares criollos.
Figura 2: Concentración de fibra detergente neutro (FDN) del rastrojo de tres cultivares de maíz sin colonizar y colonizado con Ganoderma lucidum
La barra es la diferencia honestamente significativa para la comparación entre medias.
abc Cifras con las mismas letras, son iguales (P≤0.05).
Los rastrojos colonizados con G. lucidum, mantuvieron concentraciones altas de FDN (P<0.05) en comparación a los demás tratamientos. Para estos tratamientos, el criollo A presentó la menor concentración (68.6 %) que los cultivares B Aspros® y criollo C que fueron iguales (P>0.05) entre ellos. Al comparar, la colonización con G. lucidum con los respectivos rastrojos en su forma natural, la concentración de FDN fue mayor (P<0.05) en 8.6, 3.2 y 8.1 unidades porcentuales para los cultivares criollo A, B Aspros® y criollo C, respectivamente. En el cultivar B Aspros® el rastrojo colonizado fue similar en concentración de FDN a los demás rastrojos que fueron esterilizados; el criollo C y el rastrojo colonizado fue igual en concentración de FDN a su respectivo tratamiento de esterilizado, pero fue diferente al rastrojo humedecido por 15 días en la caja Petri (P<0.05); mientras que para el criollo A el tratamiento de rastrojo colonizado fue igual en concentración de FDN al rastrojo esterilizado y mantenido húmedo por 15 días y diferente al rastrojo esterilizado y secado (Figura 2). Ganoderma lucidum en su estado puro mostró ser muy (P<0.05) fibroso, al encontrase valores de FDN que alcanzaron 88.2 % (Figura 2).
La concentración de FDA en los cultivares fue diferente (P<0.0001), con un efecto mayor (P<0.0001) del acondicionamiento a la que se sujetó el rastrojo (Figura 3), además de un efecto de interacción (P<0.0001). Los rastrojos sin esterilizar tuvieron concentraciones de FDA similares y fueron las más bajas (29.0 a 29.9 %) comparadas a los otros tratamientos. El rastrojo esterilizado y secado el mismo día tuvo mayor concentración (P<0.05) de FDA que el rastrojo sin esterilizar con diferencias de 5.1, 2.8 y 5.7 unidades porcentuales para los cultivares criollo A, B Aspros 1503® y criollo C, respectivamente. Entre los cultivares, sólo se encontró diferencia entre el rastrojo esterilizado del criollo C y el B Aspros® con tres unidades porcentuales.
El rastrojo esterilizado y mantenido húmedo por 15 días tuvo también mayor concentración (P<0.05) de FDA, en comparación al rastrojo sin tratar en proporciones de 2.9, 2.2 y 4.4 unidades porcentuales en los cultivares criollo A, B Aspros 1503® y criollo C, respectivamente. Al comparar este tratamiento de rastrojo con el esterilizado sólo se detectó diferencia (P<0.05) en el criollo A de 2.2 unidades porcentuales, los otros dos cultivares fueron iguales. Así mismo, entre cultivares para este tratamiento sólo existió diferencia entre los dos criollos, donde el C tuvo 2.4 unidades porcentuales más que el A (Figura 3).
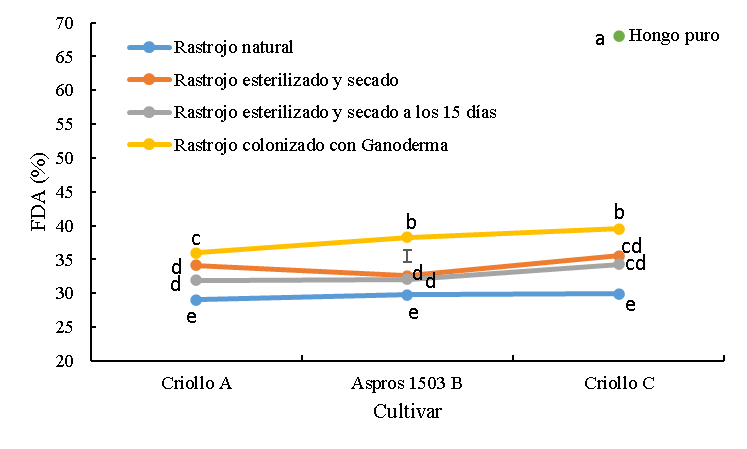
Figura 3: Concentración de fibra detergente ácido (FDA) del rastrojo de tres cultivares de maíz sin colonizar y colonizado con Ganoderma lucidum
La barra es la diferencia honestamente significativa para la comparación entre medias.
abc Cifras con las mismas letras, son iguales (P≤0.05).
El rastrojo colonizado con G. lucidum tuvo mayor concentración (P<0.05) de FDA comparado con el rastrojo sin tratar en el orden de 7.0, 8.5 y 9.6 para los cultivares criollo A, B Aspros® y criollo C, respectivamente. Al comparar este tratamiento con el rastrojo esterilizado las diferencias fueron de 1.9, 5.7 y 3.9 unidades porcentuales para los cultivares criollo A, B Aspros® y criollo C, respectivamente. Las diferencias en concentración de FDA para el rastrojo colonizado y el rastrojo esterilizado y mantenido húmedo por 15 días fueron de 4.1, 6.3 y 5.2 unidades porcentuales para los cultivares criollo A, B Aspros® y criollo C, respectivamente. Entre cultivares dentro de este tratamiento sólo se encontró diferencia (P<0.05) en la concentración de FDA para el criollo C con el Criollo A por 3.5 unidades porcentuales. El hongo G. lucidum en su estado puro tuvo la concentración mayor (P<0.05) de FDA respecto a los demás tratamientos alcanzando un valor de 68 % (Figura 3).
La concentración de lignina en los cultivares fue diferente (P<0.0001), con un efecto considerable (P<0.0001) del acondicionamiento al que estuvieron sometidos los rastrojos y una interacción, aunque ligera, pero significativa (P<0.0001) como se muestra en la Figura 4. Los rastrojos de los cultivares de maíz en su estado natural mostraron los valores más bajos (P<0.05) de lignina (2.1-2.5 %); el criollo C fue diferente (P<0.05) del B Aspros® por 0.4 unidades porcentuales (Figura 3).
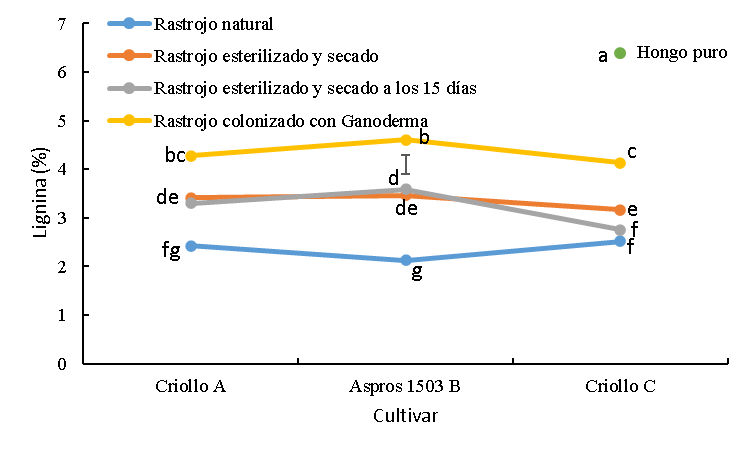
Figura 4: Concentración de lignina en tres cultivares de maíz sin colonizar y colonizado con Ganoderma lucidum
La barra es la diferencia honestamente significativa para la comparación entre medias.
abc Cifras con las mismas letras, son iguales (P≤0.05).
El rastrojo esterilizado tuvo mayor (p<0.05) concentración de lignina que el sin tratar en el orden de 1.0, 1.4 y 0.7 unidades porcentuales para los cultivares criollo A, B Aspros 1503® y criollo C, respectivamente; dentro de cultivares para este tratamiento no hubo diferencias (Figura 4).
Para el tratamiento de esterilizado y mantenido el rastrojo humedecido por 15 días, la concentración de lignina fue mayor para el tratamiento sin esterilizar en el orden de 0.9, 1.5 para los cultivares criollo A y B Aspros®, excepto en el criollo C donde se encontró mayor concentración de lignina para el tratamiento esterilizado por 0.4 unidades porcentuales. El criollo C resultó con menor concentración de lignina en el tratamiento de esterilizado y humedecido por 15 días a los cultivares criollo A y B Aspros® por 0.5 y 0.8 unidades porcentuales (Figura 4).
La concentración de lignina para el rastrojo colonizado con G. lucidum de los tres cultivares fue mayor (P<0.05) que para los otros tratamientos. Las diferencias contra el rastrojo sin tratar fueron del orden de 1.9, 2.5 y 1.6 unidades porcentuales para los cultivares criollo A, B Aspros® y criollo C, respectivamente. Con relación a los tratamientos esterilizados sin inoculación, el rastrojo colonizado tuvo mayor concentración de lignina alrededor de una unidad porcentual para los cultivares criollo A y B Aspros®, mientras que para los mismos tratamientos del rastrojo del criollo C la diferencia fue de 0.9 y 1.3 unidades porcentuales más para el colonizado. Dentro de este tratamiento con G. lucidum, el Criollo C tuvo menor concentración de lignina que el B Aspros® por 0.5 unidades porcentuales. El hongo en su estado puro alcanzó la concentración de lignina más alta con 6.4 % (Figura 4).
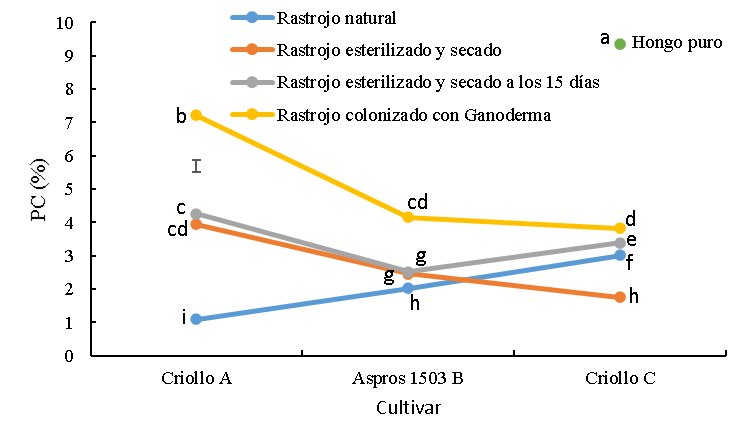
Los cultivares fueron diferentes (P<0.0001) en PC; se tuvo un efecto mayor (P<0.0001) sobre esta variable debido acondicionamiento, así como, un efecto de interacción (P<0.0001) como se muestra en la Figura 5. En el rastrojo sin esterilizar se encontraron las menores (P<0.05) concentraciones de PC, específicamente para los cultivares criollo A y B Aspros® con 1.1 y 2.0 %, respectivamente. El criollo C fue superior (P<0.05) a los dos cultivares referidos, alcanzando 3 % de PC. Con la excepción del cultivar criollo C, los rastrojos sometidos a esterilización tuvieron mayores (P<0.05) concentraciones de PC que sus respectivos rastrojos sin tratar. Así mismo, los rastrojos colonizados con G. lucidum mostraron mayor (P<0.05) concentración de PC que los rastrojos sin inocular y esterilizados alcanzando valores de 7.2, 4.2 y 3.8 % para el criollo A, B Aspros® y criollo C, respectivamente. Ganoderma lucidum en su estado puro tuvo la concentración mayor (P<0.05) respecto a todos los demás tratamientos alcanzando un valor de 9.4 % (Figura 5).
Figura 5: Proteína cruda (PC) del rastrojo de tres cultivares de maíz sin colonizar y colonizado con Ganoderma lucidum
La barra es la diferencia honestamente significativa para la comparación entre medias.
abc Cifras con las mismas letras, son iguales (P≤0.05).
Discusión
Los rastrojos esterilizados, a diferencia del que no fue tratado de esta manera, tuvieron pérdida de sustancias solubles en agua. En varios estudios se han registrado pérdidas de carbohidratos, proteínas solubles, ácidos orgánicos e inorgánicos y minerales cuando se esteriliza con vapor a altas presiones(5,6). Esta situación explica el porqué de la presencia de menor concentración de fibras en los rastrojos sin esterilizar, pues al no tener pérdidas de nutrientes por la lixiviación que ocurre, su contenido de substancias solubles (principalmente en lo que resta del contenido celular) en términos de peso, fue mayor. Se da, por tanto, un efecto de dilución en la determinación de concentración del FDN en los rastrojos que fueron esterilizados en la autoclave.
A nivel de cultivar, se observa que estos son diferentes en la cantidad de paredes celulares y contenido celular. El criollo A presentó menor cantidad de pared celular (ya sea por paredes más delgadas o diferente composición química) que los otros dos cultivares, donde el híbrido se mostró más fibroso, distinción posiblemente genética relacionada con la arquitectura de la planta la cual contrastó con los criollos por ser de hojas erectas. La característica de hojas erectas es genética y se ha incluido en los híbridos para aumentar densidades, captar más radiación solar y por tanto aumentar rendimiento(22,23). Dicha característica implica un aumento en la nervadura de la hoja con cambio en el patrón de venación y esclerénquima(24) que puede resultar en mayor concentración de fibras. Diferencias en la concentración de fibras de la pared celular entre cultivares, entre ellos varios híbridos, han sido encontrados por varios autores, lo que constata una diversidad(15,25) entre cultivares.
Los rastrojos colonizados con G. lucidum resultaron ser más fibrosos que los rastrojos sin esterilizar como lo demostró su contenido de FDN. Así mismo, el hongo resultó ser muy fibroso como lo constató su resultado. Varios estudios reportan que en G. lucidum se sintetizan compuestos fibrosos en el micelio en concentraciones elevadas de varios polisacáridos como la manosa, xilosa, arabinosa, galactosa, glucosa y ramnosa(9-12) además de quitina(13). Por tanto, en el rastrojo colonizado se tuvo a los compuestos insolubles en el detergente neutro del hongo y las fibras del rastrojo, dando como resultado un incremento del FDN. Se esperaba que se hubiera tenido cierta degradación de la pared celular por el hongo, pero es posible que el tiempo que se tuvo de colonización y desarrollo micelial no haya sido el suficiente para que se diera un efecto significativo en la solubilización de lignina. Se ha observado que G. lucidum, en una mezcla de maple, castaño y mora con otros ingredientes, comienza a degradar lignina de tres a cinco meses después de la inoculación(8) y en este estudio con rastrojo de maíz sólo se tuvieron 15 días bajo colonización, por lo que se infiere que el tiempo pudo no ser suficiente para detectarse un efecto en los componentes de la pared celular. Aparentemente, el hongo se alimentó primeramente del contenido celular del rastrojo y por eso es que no se vio efecto significativo a la baja en la concentración de FDN. En rastrojos de diversos híbridos de maíz inoculados con los hongos como Sporotrichum pulverulentum, Bjerkandera adusta y Trametes trogii(15), se presentan mayores concentraciones de las enzimas β-glucosidasa y exoglucanasa en periodos de incubación de 15 días. Ello indica que hay preferencia por las partes que contienen azúcares reductores, entre ellos los que se encuentran más fácilmente disponibles en el contenido celular. Varios hongos, entre ellos los de pudrición café (Serpula lacrymans, Coniophora puteana y Gloeophyllum trabeum, entre otros) consumen grandes cantidades de azúcares fermentables(26), los cuales se encuentran fácilmente disponibles en el contenido celular. Lo mismo ocurre con algunos otros hongos de la pudrición blanca que consumen azúcares no-estructurales en mayor proporción en el inicio de la colonización(27).
La comparación de los resultados aquí obtenidos con G. lucidum, con los de otras investigaciones, resulta un tanto difícil. Esto se debe a que los substratos utilizados estuvieron compuestos de 80 % de rastrojo de maíz mezclado con otros ingredientes derivados de granos(28). Así mismo, no se mencionan los tiempos de crecimiento del hongo; pero han encontrado disminuciones en FDN de cinco unidades porcentuales en el substrato inoculado (59.76 %) respecto al testigo sin inocular (64.94 %)(28), lo cual contrasta con los aquí encontrados.
La FDN fue la principal diferencia entre cultivares, puesto que en FDA y lignina (Figura 3 y 4) estos tuvieron concentraciones similares. Por tanto, las diferencias primarias entre los cultivares se debió a la concentración de las hemicelulosas y demás sustancias solubles en el detergente neutro. La tendencia de aumento en la cantidad de fibra en los rastrojos tratados (esterilizados) se mantuvo, así también fue para los rastrojos que tuvieron la presencia del hongo, los cuales, al tener mayor cantidad de material insoluble en FDA y en lignina, mostraron los valores más altos.
Los valores de FDA, en el criollo A fueron menores al rastrojo secado después de la esterilización. Esto implica que ocurrió un cierto efecto de hidrólisis sobre la pared celular, lo que contribuyó a que se manifestara la interacción presentada. En el rastrojo del criollo C se tuvo mayor concentración de lignina, y no mostró cambios significativos cuando se esterilizó y se secó a los 15 días, aspecto que contribuyó a la manifestación del efecto de interacción. Este comportamiento sugiere que existen diferencias en la composición de la pared celular entre cultivares.
Los rastrojos colonizados con Ganoderma lucidum mostraron las mayores concentraciones de FDA y lignina. Aun cuando algunos hongos comestibles tienen la capacidad de degradar por medio de enzimas y otros compuestos, la lignina de los forrajes(2), se observó que, en el rastrojo de maíz colonizado con G. lucidum, tal efecto no ocurrió en la extensión que se esperaba. Esto puede ser atribuible a que este hongo contiene quitina(13) y trazas de lignina(9-12) por lo que, los rastrojos colonizados con este hongo mostraron mayor concentración en estas dos variables, dándose así un efecto de aumento. Una respuesta opuesta se ha encontrado en rastrojo de maíz con contenidos de lignina de 14.9 % inoculado con G. lucidum(29). Este hongo redujo seis unidades porcentuales la concentración de lignina, aunque no se especifican los tiempos de crecimiento del hongo ni la forma de preparación del rastrojo.
Los rastrojos esterilizados y secados el mismo día, así como los esterilizados y guardados 15 días en cajas Petri, fueron los más digestibles en comparación a los rastrojos sin esterilizar y a los colonizados con G. lucidum. Se observa que, aunque estos tratamientos tuvieron mayor concentración de las tres fibras, la presión, la temperatura y posiblemente el vapor presentado en la esterilización, tuvieron un efecto en el aumento de la digestibilidad. Se ha observado que el rastrojo de maíz tratado con vapor presurizado mejora la digestibilidad por reducciones de la polimerización, ruptura de los enlaces entre la hemicelulosa, celulosa y lignina, hidrolización de hemicelulosa, incremento de la porosidad y cambios en la estructura cristalina de la celulosa(30).
Hubo también un efecto mayor de hidrólisis en el criollo C cuando se mantuvo al rastrojo humedecido por los 15 días comparado al que se secó inmediatamente después de esterilizar, aspecto que se contribuyó a la manifestación de la interacción. Esto posiblemente sucedió por diferencias en la composición de los compuestos de la hemicelulosa que se sintetizan. Los sustratos al ser esterilizados a temperaturas mayores a los 100 °C, con tiempos de duración del tratamiento térmico por más de 45 minutos y mantenidos húmedos, pierden estructura y función de sus macromoléculas por efecto de desnaturalización e hidrólisis(31,32). Las temperaturas mayores de 85 °C rompen parcialmente puentes de hidrógeno del complejo lignina-celulosa solubilizando los azúcares simples(33), por lo que puede ser la explicación del aumento de la digestibilidad en estos tratamientos.
En su estado natural, el rastrojo del híbrido Aspros 1503® y del criollo C fueron menos digestibles que el criollo A, por lo que queda de manifiesto que existen diferencias en digestibilidad entre cultivares(15). Asimismo, los valores más bajos de digestibilidad se observaron en los tratamientos colonizados con G. lucidum, situación en la que influyó la presencia de los componentes del micelio del hongo como quitina, lignina y polisacáridos estructurales(9-12), el cual resulta ser poco digerible.
El contenido de PC en el rastrojo de maíz de los diferentes cultivares se vio favorecido al ser colonizado con G. lucidum. Del micelio de este hongo se han aislado varias proteínas, como la LZ-8 la cual contiene 110 residuos de aminoácidos(34). Además, se han encontrado un complejo polisacárido-proteína que contiene una cantidad diversa de aminoácidos esenciales(35). Este contenido de aminoácidos posiblemente permitió elevar el contenido proteico del rastrojo. En algunas otras especies de hongos, específicamente en cepas de Ceriporiopsis subvermispora, Lentinula edodes, Pleurotus eryngii o Pleurotus ostreatus, se ha encontrado que elevan el contenido aproximadamente 30 % de proteína cruda de los sustratos(6), efecto manifestado mayormente para el Criollo A.
Respecto a los rastrojos esterilizados secados al momento después de la esterilización y los mantenidos húmedos en caja Petri por 15 días, estos tuvieron mayor concentración de PC respecto a los rastrojos sin esterilizar. Esto probablemente ocurrió debido a un efecto de hidrólisis en los compuestos de las paredes celulares, dado que en los sustratos esterilizados a temperaturas arriba de 100 °C y con humedad, las macromoléculas pierden estructura y función(31,32). El rastrojo criollo C esterilizado y mantenido húmedo a cero días fue la excepción, el cual por alguna razón disminuyó su contenido de proteína cruda, posiblemente más soluble, lo que contribuyó a su pérdida, contribuyendo al efecto de interacción encontrada. Al respecto se requerirán de más estudios al respecto para determinar las causas.
Conclusiones e implicaciones
Los rastrojos en su estado natural fueron diferentes en la concentración de FDN y proteína cruda, mientras que en FDA y lignina fueron similares. Por su parte, la DIVMS fue diferente, siendo el Criollo A el más digestible. La esterilización con vapor incrementó en los rastrojos la concentración de FDN y FDA. Este efecto también se notó en la concentración de lignina, con excepción del criollo C que tuvo mayor interacción negativa. La esterilización afectó diferentemente la concentración de proteína cruda en los genotipos de maíz, aumentando en uno, permaneciendo iguales en otros y disminuyendo en uno más. Así mismo, la esterilización con vapor indujo valores mayores de digestibilidad, indicando un efecto fuerte de la alta temperatura y presión, principalmente. La colonización con el hongo G. lucidum durante 15 días no mejoró la digestibilidad in vitro, pues la afectó negativamente al incrementar las concentraciones de fibras, manifestándose en valores menores de digestibilidad. El único parámetro que se mejoró en los rastrojos colonizados con G. lucidum fue la concentración de proteína cruda, mostrando mayor concentración que sus respectivos rastrojos sin inocular y esterilizados, alcanzando valores de 7.2, 4.2 y 3.8 unidades porcentuales para el criollo A, B Aspros® y criollo C, respectivamente. Los resultados aquí obtenidos evidencian que se requieren mayores tiempos de crecimiento de G. lucidum en el sustrato de rastrojo de maíz para detectar cambios en la pared celular, principalmente por solubilización de lignina y otros elementos que se encuentran en esta estructura para que esto se manifieste en mejoras de la digestibilidad.
Agradecimientos
La primera autora agradece la beca otorgada por el Consejo Nacional de Ciencia y Tecnología para realizar estudios doctorales.
Literatura citada: