https://doi.org/10.22319/rmcp.v13i2.5922
Review
Demi-embryo reconstitution, a factor to consider for the success of embryo bisection. Review
Alfredo Lorenzo-Torres a
Raymundo Rangel-Santos a*
Agustín Ruíz-Flores a
Demetrio Alonso Ambríz-García b
a Universidad Autónoma Chapingo, Posgrado en Producción Animal. Carretera México-Texcoco, Km 38.5, 56230. Texcoco, Estado de México, México.
b Universidad Autónoma Metropolitana, Departamento de Biología de la Reproducción, Ciudad de México, México.
* Corresponding author: rangelsr@correo.chapingo.mx
Abstract:
For many years it has been sought to increase the reproductive efficiency of livestock using biotechnologies such as embryo bisection. However, despite its potential in livestock, its level of adoption is limited. The present work reviews the importance of demi-embryo reconstitution, after bisection, and the main factors that limit its success in livestock. It is possible to increase its level of adoption if it is possible to increase the efficiency currently obtained with this technique, this can be achieved by making a more precise selection of the embryos subjected to bisection. Embryo quality is one of the most important factors related to the potential to reconstitute into viable demi-embryos after bisection, which can be used with greater reliability in embryo transfer programs.
Key words: Embryo bisection, Demi-embryo reconstitution, Embryo development.
Received: 07/01/2021
Accepted: 15/07/2021
Introduction
Embryo bisection is a reproductive biotechnology that allows obtaining identical demi-embryos, to be used in research(1) or the livestock industry(2). The aim of embryo bisection is to increase the number of semi-embryos available for transfer and therefore, the offspring of genetically superior animals(1,3,4). This technique can be applied to embryos developed in the morula or blastocyst stage and consists of obtaining two similar halves by mechanical bisection(5,6,7). Morula bisection can be carried out in any position of the embryo, due to its symmetric morphology(8). In the case of blastocysts is important the symmetrical orientation of the embryo to obtain a proportional distribution of the inner cell mass (ICM) and trophectoderm (TE) in the resulting demi-embryos(9).
Embryo bisection is performed in embryos of several species(10,11,12), in order to increase embryo availability(12), pregnancy rate(13), and the number of offspring(5,14,15). However, there are studies where the survival of demi-embryos was low(16,17), even lower compared to the use of whole embryos(18). This could be associated with the fact that embryo bisection is an invasive technique(6) and the procedure causes cellular damage(19). Therefore, the success of the technique could be influenced by factors associated with the original embryo(20,21,22) and its ability to reconstitute itself in the resulting demi-embryos(23). The objective of this review is to highlight the importance of demi-embryo reconstitution in embryo bisection programs, as well as to discuss the main factors that influence its success.
Importance of embryo bisection in livestock
Embryo bisection has been carried out in different livestock species of interest, such as rabbits(19), sheep(24), bovines(12), goats(10), equines(25), swine(26), and even in humans(22). Despite it is an invasive technique, it is practical and it does not require cell reprogramming like cloning(6). Embryo bisection in livestock allows to produce identical twins for experimental use(14), reducing the number of animals needed per treatment for comparison tests(27) or to increase the availability of transferred embryos(28). In addition, obtaining identical twins facilitates the evaluation of sires or maternal trait tests(29) and, lastly, to maintain desirable characteristics in cattle(3).
Embryo bisection has allowed to increase pregnancy rate(29) and the number of offspring(13), compared to the transfer of whole embryos(13,24). Most authors have reported a higher number of semi-embryos available for transfer in relation to the number of bisected embryos, thus increasing the efficiency in the number of offspring (Table 1). However, there is a great variation (75 to 118 %, efficiency), which could be mainly associated with factors related to the original embryo. In ewes, the pregnancy rate was 64 % when they received airs of demi-embryos, obtaining 118 % efficiency in offspring(14). Likewise, there was a higher percentage of embryo survival when transferring two demi-embryos per recipient ewe (101 %, 710/705), compared to the transfer of two whole embryos (62 %, 771/1252) considering the number of original embryos(24). Further, 30 % more lambs were born after the transfer of pairs of demi-embryos, compared with whole embryos (85 vs 55 %, P<0.05)(30).
Table 1: Efficiency of embryo bisection in the production of offspring in relation to the number of bisected embryos
Species | Bisected embryos | Number of offspring born | Efficiency (%) | Reference |
Bovine | 36 | 27 | 75 | 13 |
Bovine | 50 | 61* | 105 | 15 |
Bovine | 11 | 12* | 109 | 5 |
Sheep | 40 | 34 | 85 | 30 |
Sheep | 24 | 21** | 88 | 16 |
Sheep | 705 | 710* | 101 | 24 |
Sheep | 16 | 17 | 106 | 31 |
Sheep | 39 | 46 | 118 | 14 |
* Number of fetuses diagnosed by ultrasound between 30 and 80 d of gestation or ** by sacrifice surgery after slaughter. Efficiency (%)= Number of offspring born / embryos bisected.
On the other hand, in some studies, low efficiency was obtained with embryo bisection (16,32,33). It has been reported that a lower percentage of lambs were born after the transfer of bisected sheep embryos, compared to whole embryos (27 vs 52 %, P<0.05)(34). However, in beef cattle, despite the problems being associated with twin gestation(4), implementing embryo bisection represents an economic benefit once the number of offspring is increased(2,17,35). Therefore, embryo bisection can be implemented in embryo transfer programs(2,24).
Embryonic reconstitution after bisection
Embryo reconstitution is an indicator of the ability of demi-embryos to become offspring after their transfer to a recipient female(18,29). In adult tissues, stem cells are responsible for repairing injuries and regenerating tissues(23,36) due to aging or diseases(37). In the case of embryos, something similar happens, through the replacement of specialized cells that have been lost due to some alteration(23). Embryos are capable of repairing their injuries, adapting to environmental conditions to survive after their reconstitution(23). Embryonic cells in early stages can adapt both in mitotic rate and differentiation process(38), due to the plasticity they present(39,40). In addition, it has been proven that embryonic cell conglomerates in the early stages of development can become living organisms through cell reorganization, such as the case of blastocyst splitting(41).
Thus, a group of cells has properties that exceed the potency of any of the individual cells within the group for cell reconstitution, which could be a joint effect(23). The extruded cells or even cell debris observed after embryo bisection could contain enough viable cells to proliferate and reorganize, originating another functional embryo(8). At the time of embryo bisection, the resulting halves can be cultured in vitro for 2 to 48 h(5,26). In each demi-embryo, the ICM is reorganized and the reconstitution of the blastocele begins immediately(5,29,42). The union of the edges of trophoblastic cells during bisection is responsible for the trophoblast's ability to reconstitute itself(5) since this group of cells secretes fluid into the blastocele, a process regulated by genes(43,44). This allows the spherical formation of demi-embryos(42), being the trophoblastic cells important for embryo implantation(45). Likewise, an embryo, even if it has lost half of its cells, it is still an organism and a characteristic of organisms is to repair their lesions, regenerating them to continue their development(23).
In the laboratory, it was bisected sheep expanded blastocysts produced in vitro with a microblade (Figure 1a), using the procedure called scratched bottom dish(46) (Figure 1b) and it was observed a completely spherical reconstitution (70 % the size of the original embryo) 12 h after in vitro culture (Figure 1c).
Figure 1: Embryo bisection process and reconstitution of the demi-embryos after 12 h of in vitro culture
a) Expanded blastocyst oriented symmetrically with respect to the bisecting microblade, b) resulting demi-embryos, and c) reconstituted demi-embryos. ×200.
Several authors have reported a reconstitution percentage ranging from 90 to 178 % (Table 2).
Table 2: Reconstitution efficiency of bovine demi-embryos after in vitro culture for 2-48 h post-bisection
Bisected embryos | Number of demi-embryos | Reconstitution* (%) | Reference |
21 | 19 | 90 | 29 |
11 | 16 | 145 | 5 |
176 | 268 | 152 | 12 |
19 | 30 | 158 | 47 |
230 | 408 | 178 | 28 |
*Reconstitution (%) = Number of demi-embryos / bisected embryos.
The evaluation of embryo reconstitution could allow the selection of viable demi-embryos and it can be a useful tool in embryo transfer programs(18).
Factors that affect demi-embryo reconstitution
Embryo bisection technique
Embryo bisection is a technique that allows the production of identical twins in embryo transfer programs(5). In the 80s, the technique required up to six embryo manipulation and bisection instruments(5). However, over time, different methodologies have been developed to simplify the technique(1), since the procedure required up to 15 min to bisect an embryo(48). In addition, the use of cutting instruments, such as microblade(5,25,49) or glass needle(50,51,52), have been extensively studied, with the objective of minimizing cellular damage at the time of cutting(53). Thus, the success of the technique depends on the minimum damage produced to the embryos(19), since the procedure generates 10 to 13% of cell loss(47,54).
On this regard, the microblade embryo bisection method has proven to be practical and with an application under field conditions(1). The implementation of this technique has been simplified by means of vertical pressure at the time of embryo bisection, using a microblade adapted to a single micromanipulator, without the use of an embryo holding micropipette(12,15,34). In the laboratory, has been observed that the use of the scratched bottom dish technique(46) with 50 µL of a commercial bisecting medium, facilitates embryonic fixation and prevents cell adhesion to bisecting and culture materials. This makes it possible to bisect groups of five embryos in approximately 3 min, making the application of this biotechnology more practical and without subjecting the embryos to prolonged stress.
On the other hand, there is evidence showing that the technician who performs the embryo bisection influences the productive response. In sheep, it has been evaluated the effect of the technician at the time of bisection, finding a significant difference between the two technicians in pregnancy rate (66 vs 75 %, P<0.05) and demi-embryo survival (51 vs 44 %, P<0.01)(55). Thus, proper training of the technician should be considered before implementing embryonic bisection.
Developmental stage
The stage of embryonic development, morula, early blastocyst, or expanded blastocyst at the time of bisection, is one of the most important factors that affect the pregnancy rate of transferred embryos(8). After the bisection of mouse embryos, it was found that, in the morula stage, the demi-embryos are reconstituted in a lower percentage than in the blastocyst stage (74 vs 90 %, P=0.001) after in vitro culture for 24 h(45). In cattle, no significant differences were reported when transferring demi-embryos from morulae, early blastocysts, and expanded blastocysts, on pregnancy rate (51-65 %, P>0.1)(53). In another study, there was a lower pregnancy rate using bisected morulae (7/44, 16 %) compared to early blastocysts (58/96, 60 %), P <0.01(8). Likewise, it has been reported a higher percentage of viable fetuses after blastocyst bisection compared to morulae, 91 (10/11) vs 30 % (3/10), P<0.05, at d 70 of gestation(47). Finally, in sheep, six pairs of identical twins were obtained from blastocyst bisection, without success in morula bisection (P<0.05)(16).
In a practical way, it appears that bisecting morulae is easier due to the morphological symmetry they present, however, in the laboratory, it was observed that bisecting expanded blastocysts and even hatched blastocysts was easier once the ICM and TE were clearly identified. Furthermore, some authors have reported a higher pregnancy rate using blastocysts compared to morulae (Table 3), perhaps because they are more tolerant to manipulation and are less affected by the loss of the zona pellucida(8,54). This could be due to the fact that embryogenesis is strictly regulated in time(22) and the more developed the embryos are, the more tolerant they are.
Table 3: Effect of the developmental stage of the whole embryo on the pregnancy rate of transferred semi-embryos
Species | Developmental stage, % (n) | Reference | ||
Morula | Blastocyst | Expanded blastocyst | ||
Caprine | 0 (5) | 33 (9) | 55 (11)* | 10 |
Ovine | 60 (20) | 88 (24) | - | 16 |
Bovine | 48 (162) | 60 (96) | 54 (28) | 8 |
Bovine | 51 (71) | 64 (61) | 58 (12) | 53 |
Bovine | 39 (139) | 36 (33) | 30 (10) | 12 |
%= Pregnancy rate; n= Number of recipient females; *Hatched blastocyst.
Embryo quality
There is wide evidence for the use of excellent quality embryos for bisection purposes(12,40,56,57). The embryos selected for bisection must meet certain morphological criteria, from which the success of demi-embryo reconstitution will depend(28) and, consequently, the pregnancy rate(12,51). The quality of the embryos must be excellent or good, depending on the morphological criteria(58) because when they are of low quality (fair and bad) are more vulnerable to the bisection process(12,47).
In bovines, when bisecting morulae, a higher percentage of survival was obtained in the group of excellent and good quality, compared to morulae of regular and poor quality, 167 (20/12) vs 75% (9/12), P<0.001(47). On the other hand, 42 % pregnancy was found in cows after thawing and transferring excellent quality demi-embryos, while when transferring demi-embryos from low-quality embryos, no female became pregnant(51). Likewise, it has been reported a higher percentage of development in pairs of demi-embryos, when bisecting bovine embryos of excellent quality, compared to those of good quality (76 vs 40 %, P<0.05)(12). Therefore, evaluation of embryo quality subjected to bisection should be considered to obtain positive results. However, the morphological evaluation of embryos subjected to bisection is a subjective aspect, based on the experience of the researcher.
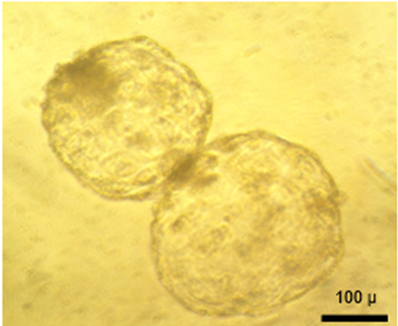
In the experience in manipulating sheep embryos of this research team, was found a higher percentage of demi-embryo reconstitution (145 %, approximately), when bisecting embryos of excellent quality and with a diameter greater than 230 µm. In the resulting demi-embryos, after 12 h of in vitro culture, was found that when they were reconstituted, they had an average diameter of 176 ± 10.03 µm (Figure 2), similar to that reported in quality two porcine demi-embryos (161.6 ± 25.7 µm)(26), but higher than that reported in human demi-embryos (121 µm)(22). Therefore, the embryonic diameter has been proposed as an indicator of quality(25,59) since the size of the embryo is important in maternal recognition(60).
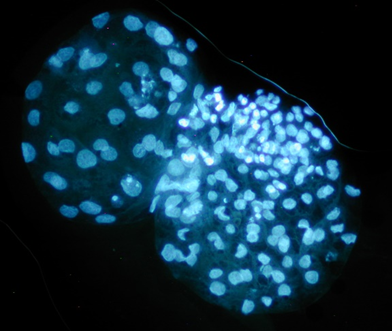
On the other hand, the size of the embryo is also associated with the number of cells(61) and, consequently, with the embryo quality. In poor quality embryos, there is a deficiency in the rate of cell division, mitosis, and consequently in the number of cells(47). Thus, the size of the embryo is proportional to the number of cells used for its reconstitution(22). There is a 50 % recovery of cells from the original embryos in the resulting demi-embryos depending on the quality and uniformity of the bisection process(26,28). This suggests that bisecting bigger size embryos will produce semi-embryos with more ICM and TE cells, thus increasing their survival. In the laboratory, was obtained an average of 68 ± 11.3 cells in reconstituted demi-embryos, after 12 h of in vitro culture (Figure 3), from embryos with 122 ± 6.6 cells. In this sense, active cell proliferation can be a criterion of embryonic quality(62). Based on the above, the diameter of the embryo could be an objective criterion to select embryos for bisection, in order to achieve the greatest possible success in demi-embryos reconstitution.
Figure 2. In vitro reconstitution of demi-embryos after 12 h of culture. ×200
Figure 3. Cell staining (Hoechst) of in vitro demi-embryos after 12 h of culture. ×200
In vitro or in vivo produced embryos
There are differences between embryos produced in vitro or in vivo, both in morphology and in molecular components(63), where in vivo produced embryos are of better quality. Nevertheless, a high percentage of in vitro survival of ovine demi-embryos has been reported after bisection (80-85 %), and 33 % pregnancy after transferring pairs of demi-embryos to recipient ewes (5/15)(34). On the other hand, for embryos produced in vivo, some authors reported greater pregnancy rates. In bisected bovine embryos produced in vivo and cultured the demi-embryos in vitro, a high percentage of reconstitution was reported (47/60 , 78.3 %)(15). In bovines, there was 53 % (18/34) pregnancy when transferring embryos produced in vivo, bisected, and cultured in vitro(18). Thus, the efficiency of embryo bisection in relation to the origin of the embryo seems to be lower in embryos from in vitro conditions. This could be due to the low quality and efficiency of in vitro production(63). Therefore, it is necessary to improve efficiency in both embryo production procedures, once the two of them are focused on improving productivity in livestock production(64).
Effect of breed and age of embryo donors for bisection
There are other little-studied factors that could affect the embryo's fate under the bisection process. In sheep, the effect of the breed was evaluated, without finding significant differences in pregnancy rate and survival of demi-embryos between embryos from Gotland and Finnish Texel (69 vs 50 % and 42 vs 26 %, respectively, P>0.05) and between Danish Texel and Finnish Texel breeds (74 vs 74 % and 50 vs 50 %, respectively, P>0.05)(55). In addition, the effect of the donor age has been reported, without finding differences in pregnancy rate between embryos generated from adult females (24 months of age) and young (approximately 10 mo of age) (74 vs 74 %, P>0.05), however, demi-embryo survival (51 vs 47, P<0.05) and percentage of identical twins was higher in adult ewes than in young (38 vs 27, P<0.01)(55). This could be related to a lower survival capacity of whole embryos from young ewes(65,66,67), which is confirmed after transferring demi-embryos(55).
Conclusions
Demi-embryos reconstitution is a key factor for the success of embryo bisection and the highest efficiency is obtained by selecting excellent quality embryos regardless of their developmental stage. The results of the literature show the potential of the bisection technique; therefore, its application should be considered to improve the efficiency of embryo transfer programs.
Literature cited: