https://doi.org/10.22319/rmcp.v14i2.5881
Revisión bibliográfica
Importancia de Haematobia irritans en la ganadería bovina de México: Situación actual y perspectivas. Revisión
Roger Iván Rodríguez Vivas a*
Carlos Cruz Vázquez b
Consuelo Almazán c
Juan José Zárate Ramos d
a Universidad Autónoma de Yucatán. Facultad de Medicina Veterinaria y Zootecnia. Carretera Mérida-Xmatkuil Km 15.5, Mérida, Yucatán, México.
b Tecnológico Nacional de México. Instituto Tecnológico El Llano, Aguascalientes, México.
c Laboratorio de Inmunología y Vacunas; Facultad de Ciencias Naturales, Universidad Autónoma de Querétaro, Querétaro, México.
d Universidad Autónoma de Nuevo León. Facultad de Medicina Veterinaria y Zootecnia, Campus de Ciencias Agropecuarias. General Escobedo, Nuevo León, México.
Autor de correspondencia: rvivas@correo.uady.mx
Resumen:
La mosca de los cuernos Haematobia irritans es un ectoparásito hematófago cosmopolita de gran importancia en la ganadería. En México, H. irritans se distribuye por todo el país y se encuentra durante todo el año. La fluctuación de la población de H. irritans está relacionada con las condiciones climáticas. A pesar de su amplia distribución, los efectos sobre la salud animal y su impacto negativo en la producción de carne y leche, existen pocos datos sobre su infestación y su epidemiología es limitada. Este trabajo es una revisión sobre la situación actual de H. irritans en bovinos en México, su impacto económico, métodos de control, perspectivas y oportunidades de investigación.
Palabras clave: Haematobia irritans, Mosca del cuerno, Epizootiología, Control.
Recibido: 25/11/2020
Aceptado: 15/09/2022
Introducción
La mosca del cuerno, Haematobia irritans (Linneaus, 1758), es un díptero de la familia Muscidae de importancia económica en la ganadería bovina debido a sus hábitos hematófagos(1). Su importancia radica en pérdidas directas debido al consumo de sangre, a los daños a la piel y la intranquilidad constante que provoca al alimentarse, lo que ocasiona la reducción de la ganancia diaria de peso y la producción de leche(2). El impacto en la producción está relacionado con el nivel de infestación y depende de algunas características intrínsecas de los animales y de las condiciones ambientales de la región ecológica donde se desarrollan(3). En México, H. irritans se distribuye en todo el país y se ha reportado su presencia durante todo el año(2).
El control de las infestaciones por H. irritans se realiza principalmente mediante el uso de insecticidas de diferentes familias químicas tales como piretroides, organofosforados, fenilpirazolonas, reguladores del crecimiento e inhibidores del crecimiento de los insectos, entre otros; sin embargo, su uso frecuente e irracional han provocado la selección de poblaciones resistentes a los insecticidas, principalmente a los piretroides y organofosforados(2).
Alternativamente, el control de esta mosca se puede realizar de forma sustentable mediante manejo cultural del estiércol así como la aplicación de diferentes medidas de control biológico, que incluye, entre otros, la liberación de enemigos naturales, agentes entomopatógenos y el uso de sustancias de origen botánico con capacidad de repeler o matar a estos insectos; el control inmunológico, mediante la vacunación, con el fin de evitar la alimentación o disminución del consumo de sangre, es una alternativa que se ha ensayado de manera preliminar, sin embargo, a la fecha, no existen vacunas contra la infestación por esta mosca(4). El control integrado de plagas es el método más apropiado para reducir las poblaciones de moscas en el ganado y para ello se requiere un uso combinado y racional de los métodos existentes(2).
En este trabajo se presenta una revisión sobre la situación actual del parasitismo por H. irritans en la ganadería bovina, el impacto económico y las medidas de control disponibles, así como las perspectivas y oportunidades de investigación en México.
Efectos directos e indirectos que ocasiona H. irritans a los bovinos
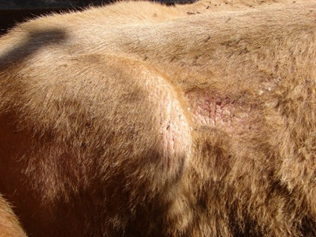
Daños directos a los animales. Las hembras y machos de H. irritans se alimentan de 20 a 38 veces al día, consumiendo pequeñas porciones de sangre, en promedio cada mosca ingiere 10 µl al día(5). Su acción hematófaga produce daños en la piel, lo que ocasiona una reducción en su calidad(3). Los defectos en la piel incluyen puntos negros y orificios, donde la mayor parte del daño parece ser el resultado de respuestas inflamatorias dérmicas (Figura 1). En el sitio de alimentación se produce infiltración eosinofílica, foliculitis eosinofílica y furuncolosis con alopecia(6).
Figura 1: Afectación severa de la piel con alopecia e hiperqueratosis en un bovino hembra sin tratamiento contra ectoparásitos durante la época de mayor intensidad de Haematobia irritans en la región norte del Golfo de México
Imagen C. Almazán
Enfermedades que transmiten. H. irritans es hospedero intermediario de Stephanofilaria stilesi, nematodo que causa lesiones cutáneas en el ganado y que ha sido reportado en el ganado del oeste y suroeste de los Estados Unidos de América (EUA) y Canadá; puede transmitir mecánicamente varias especies de bacterias del género Staphylococcus, que causan mastitis en las vacas lecheras(7). Adicionalmente, están involucradas en la transmisión mecánica de otros patógenos tales como Trypanosoma vivax y T. evansi, Francisella tularensis, Corynebacterium pseudotuberculosis, Parabronema skrjabini y Anaplasma marginale(8-10).
Impacto económico
Las pérdidas económicas debidas a la infestación por H. irritans en ganado productor de carne se han estimado en Brasil(11) en pérdidas promedio anuales de 3.25 kg/vaca y en Argentina de 0.028 kg/vaca/día (305 días de lactancia, 8.54 kg por vaca)(12). En los EUA, encontraron que las novillas que recibieron tratamiento insecticida contra la mosca del cuerno tuvieron 14 % más de ganancia de peso en comparación de las que no lo recibieron(13). El control de moscas resulta también favorable en las vacas al ganar 14.4 kg más de pesos que las vacas que no reciben control(14).
Las pérdidas en la producción de leche causada por H. irritans en explotaciones lecheras en EUA se estimaron en 27 kg de leche/vaca/año(15). Se calcula que durante el año 2016 en EUA se perdieron 1,750 millones de dólares por la acción directa de H. irritans sobre los animales; además de considerar un gasto adicional de 60 millones de dólares anuales por concepto del uso de insecticidas para su control(16). En México, de acuerdo a la población de ganado bovino en riesgo potencial, se estima que las pérdidas atribuidas a H. irritans ascienden a $231.66 millones de dólares al año(1). Sin embargo, no se han evaluado las pérdidas asociadas a las tasas de preñez y medidas de control, así como los patógenos que pueden transmitir.
Ciclo biológico de H. irritans
Se sugiere que H. irritans incluye a dos subespecies, morfológicamente similares, H. irritans irritans y H. irritans exigua (mosca del búfalo), la primera distribuida en Europa y América, y la segunda en Asia y Australia(17). En esta revisión se denominará H. irritans, a la mosca del cuerno que se encuentra en América.
H. irritans abunda en climas tropicales y subtropicales con rangos de temperatura de 20 a 30 0C y humedad relativa de 65 a 90 %(17). En México también se distribuye en climas templados(18).
La mosca adulta de H. irritans mide de 3 a 4 mm de largo y es de color gris con rayas obscuras en el tórax, posee un par de ojos compuestos de coloración rojizo oscuro; en este díptero existe dimorfismo sexual, las hembras poseen los ojos más separados y pequeños que los machos, además en estos últimos el abdomen está ligeramente replegado. H. irritans se posa sobre el animal, con dirección hacia el suelo(17).
El rango de hospederos para H. irritans es muy amplio y la mosca permanece durante largos periodos de tiempos alimentándose sobre la superficie de los hospederos. El hospedero principal es el bovino, pero también parasita a ovinos, equinos, caninos, búfalos de agua, bisontes y humanos(19). Se ha observado que el color de los animales influye en las preferencias de las moscas, los animales de color negro atraen mayor número de moscas(2). Los toros son más atractivos que los novillos o las vacas. H. irritans, permanecen la mayor parte de su vida sobre el animal e incluso realizan la cópula sobre éste. Las hembras sólo abandonan a su hospedero para depositar sus huevos en el estiércol fresco. La atracción fecal es corta y comienza a desaparecer luego de 10 min post-defecación. La permanencia de las moscas en la materia fecal también es corta y varía entre uno y 10 min(2). Durante las primeras horas del día se localiza en los hombros y dorso mientras que por la tarde las moscas se concentran en la línea media del abdomen y los costados, regresando al área de los hombros y dorso al anochecer(2). El promedio de vida de las moscas adultas es de seis a ocho semanas y esta longevidad está en relación inversa con las bajas temperaturas, que influyen en el desarrollo del ovario, el apareamiento, el desarrollo de las larvas y la emergencia del adulto(17). Posterior a la emergencia, los adultos requieren tres días para completar la maduración de los órganos reproductivos. El apareamiento ocurre de tres a cinco días después de la emergencia y la oviposición ocurre de tres a ocho días(20).
En el continente americano se ha observado que la temperatura es el principal factor climático que se asocia al ciclo de vida y está relacionado con la estacionalidad de las infestaciones por este díptero, que también se relaciona con la presentación de la diapausa facultativa de forma total o parcial. En regiones tropicales y de clima templado se ha observado menos cantidad de moscas en invierno, pero no una diapausa total, mientras que en regiones de clima templado-frío este fenómeno se presenta invariablemente en invierno(18). Se han observado poblaciones de la mosca durante todo el año en el trópico húmedo de México(21). La relación humedad/temperatura es crucial sobre la reproducción de las moscas(18).
La oviposición de H. irritans ocurre durante los primeros 2 min después de la evacuación de heces, y puede ocurrir durante el día y la noche. Una hembra puede ovipositar hasta 400 huevos, depositados en grupos de 20-25(22). Los huevos son ovales, cilíndricos, ligeramente curvados con un surco medial longitudinal y de color amarillo o blanco recién puestos para luego oscurecerse. Miden 1.0 a 0.5 mm de largo y 0.34 a 0.39 mm de ancho(17). Para que ocurra la eclosión se requieren 24 -26 ºC de temperatura y una humedad relativa cercana al 100 %. La eclosión ocurre después de 20 a 48 h de incubación(20).
Las larvas de H. irritans son de color blanco amarillento y miden 7 mm de largo, poseen un par de espiráculos posteriores que en términos generales tienen forma de “D”(20). Las larvas pasan por tres estadios denominados L1, L2 y L3. El desarrollo de L1 a L3 requiere de cuatro a ocho días y la pupación entre seis y ocho días. Las larvas L2 y L3 poseen espiráculos anteriores mientras que la L1 carece de ellos. Por su parte, el par de espiráculos posteriores permiten diferenciar entre la L2 y L3, ya que en las L2 en sus espiráculos presentan dos aberturas y en las L3 los espiráculos presentan tres. Las larvas se alimentan de las bacterias presentes en las heces(23).
El desarrollo de la pupa requiere de seis a ocho días(23). El estado de pupa está rodeado por el exoesqueleto de la L3, que se oscurece y endurece, formando una cápsula llamada pupario(23). El desarrollo de la pupa requiere condiciones de humedad y temperatura similares a las del desarrollo de las larvas. Después de siete u ocho días, los adultos emergen y buscan inmediatamente un hospedero para alimentarse(20). La diapausa ocurre a temperaturas por debajo de los 23 °C y las pupas pueden sobrevivir períodos prolongados de exposición a temperaturas tan bajas como -5 °C(22). En condiciones óptimas, el ciclo de vida se completa en 10 a 20 días(3).
Distribución geográfica y dinámica poblacional. En 1984, se notificó por primera vez la presencia de H. irritans en el estado de Veracruz. En la actualidad, se reconoce su presencia en todo el territorio nacional, y está asociada principalmente al ganado mantenido en condiciones de pastoreo, ya que esta condición facilita su ciclo de vida y mantenimiento en el ambiente(2,24).
La dinámica poblacional de H. irritans está relacionada con las condiciones climáticas regionales, y las moscas se observan durante todo el año en climas tropicales. En cada región la abundancia poblacional durante el año es diferente, pero siempre tiende a mostrar estacionalidad. En general, se pueden observar dos picos de población entre finales de primavera y principios de otoño. Además del clima regional, la abundancia también puede responder a otros factores ambientales y de manejo que pueden causar fluctuaciones poblacionales durante un año e incluso entre diferentes años(25). En sitios por arriba de los 1,800 msnm las poblaciones no se desarrollan de forma importante. En México, la dinámica poblacional de H. irritans muestra en lo general, un comportamiento bimodal, con amplias fluctuaciones a lo largo del año; se observa estacionalidad de la infestación asociada a la temperatura y humedad relativa. El índice de infestación es variable pero más importante en regiones tropicales que en templadas y con mayor altitud sobre el nivel del mar(26,27,28).
El mayor índice de infestación se puede presentar a partir de la segunda mitad de la primavera y hasta el inicio del otoño, pudiendo observarse en ciertas zonas, hasta tres picos poblacionales. El índice de infestación en el verano puede llegar a ser mayor de 4,000 moscas por animal, en tanto que en otros periodos puede variar entre 200 y 450 moscas. La aplicación de insecticidas y la resistencia a los mismos, pueden intervenir en la estimación en un hato; asimismo, este índice puede guardar relación con el manejo del pastoreo y de las excretas(28). En áreas ganaderas de clima templado, la dinámica poblacional de H. irritans es bimodal, y se considera estacional, su presencia aumenta desde el final de primavera hasta inicio del otoño, y el verano es la época con más altos índices de infestación. En estas áreas se puede presentar diapausa facultativa en el invierno lo que lleva a hacer casi imperceptible su presencia(26,27).
Se pueden producir varias generaciones de H. irritans en un año. En climas fríos se han estimado de 7 a 9 generaciones al año, mientras que en climas cálidos el número de generaciones puede oscilar entre 8 y 14(22). en una región semiárida de Brasil, se han reportado treinta generaciones por año(20). En México no existe información sobre el número de generaciones producidas por H. irritans por año. Esta información es fundamental para comprender el comportamiento del parásito y elaborar estrategias de control.
Resistencia del hospedador. Las razas de ganado Bos indicus son menos susceptibles a la infestación por ectoparásitos que las razas B. taurus(29). Asimismo, existen diferencias significativas en la densidad de H. irritans entre diferentes razas B. taurus: por ejemplo, la raza Chianina es más resistente que las razas Angus, Hereford y Charolais(30). En Brasil, se encontró que los bovinos de la cruza Guzerat x Holstein presentan mayor infestación en comparación con la raza Guzerat pura(31). En un estudio realizado en el sur de México, se comparó el ganado B. taurus con B. indicus para asociar a la raza de bovino con el factor de resistencia a la infestación y se obtuvo que los animales de la raza Brahman fueron parasitados por igual o menor número de moscas que los animales de razas B. taurus(2). La infestación por H. irritans en los animales de un hato no es homogénea y más del 50 % de la población de moscas parasitan al 15-30 % de los animales lo que sugiere que algunos bovinos son más susceptibles a la infestación(32). Los factores de susceptibilidad entre y dentro de razas de bovinos a infestaciones por H. irritans incluyen el color (los animales de color obscuro son más susceptibles), tamaño (animales de talla grande son más parasitados), densidad del pelo y producción de sebo (animales con menor densidad pelo y producción de sebo, son más parasitados), hormonas (la testosterona favorece la infestación). Además, la resistencia natural está influenciada por la respuesta inmune y el sistema de coagulación individual influencian el parasitismo(33).
Métodos para estimar infestación el número de moscas y umbral económico (UE). Para el establecimiento del UE en las infestaciones de H. irritans es fundamental determinar la cantidad de moscas presentes en los animales que causaran un daño económico. Se entiende como daño económico a la cantidad de perjuicio que justifica el costo del control artificial, y como UE a la densidad de población del parásito que se usa para tomar la decisión de iniciar una acción de control que impida una pérdida mayor al costo de la intervención(2). La determinación del número de moscas sobre los animales se realiza mediante el método visual directo (VD) o método indirecto digital (VID) a través de fotografía o video. En la mayoría de los casos se realiza a una distancia de 1 a 4 m de los animales. Eso dependerá de la docilidad de los mismos(34-36), e incluso se han descrito distancias de 5 a 10 m usando binoculares(37). Los horarios para los conteos recomendados son las horas más frescas del día, que permitan la observación con iluminación natural, además debe considerarse que en horarios más cálidos del día una alta proporción de estas moscas se desplaza hacia las partes bajas del abdomen de los animales, lo que dificulta el conteo, y éste debe realizarse siempre a la misma hora. En la mayoría de los trabajos publicados los recuentos se realizan en horarios comprendidos entre las 06:00 a las 12:00 h(16,38,39). En otros estudios, las cuantas se han realizado de 15:30 y las 19:00 h(36).
La determinación VD o VID del número de moscas por animal requiere realizarse por personal capacitado y debe realizarse de un lado del animal y posteriormente multiplicar por dos para obtener el total de moscas por animal, o bien realizarse en ambos lados por dos observadores de manera simultánea(34-36). Debe considerarse que cuando la densidad de moscas es muy alta se corre el riesgo de subestimar el número de moscas; en este sentido cuando la cantidad de moscas en las regiones escapular, inter escapular y costal son ≤25 se cuentan individualmente y cuando son ≥25 pueden contarse por grupos de 5(40).
Las regiones con mayor número de H. irritans son la región escapular, inter escapular y costal(40) (Figura 2). También pueden considerarse la espalda, flancos, las patas y ambos lados de la cabeza(35). La determinación de la infestación puede realizarse en animales confinados, semiconfinados o libres(16,34,36).
Figura 2: Infestación por Haematobia irritans en las regiones dorsal del cuello y escapular en un bovino macho de piel obscura
Imagen Dra. Ma. Lorena Torres-Rodríguez
El uso de fotografías y videos en el método VID(35,36) y videos(41), tiene las ventajas de que las imágenes captadas quedan registradas de manera permanente, las moscas visibles se cuentan con alta precisión, las imágenes son menos propensas a errores de estimación y no se requiere gran cantidad de mano de obra(36). Sin embargo, la ventaja del método VD es que es más rápido y eficiente, suficiente para medir los cambios en la densidad de población de H. irritans(39).
Para la determinación del número de moscas a ser consideradas como UE se han realizado varios estudios en diferentes partes del mundo(16,42). De esta manera se observó que 200 moscas causan la pérdida de 520 ml de leche por día y 28 g de peso vivo por animal al día(43). Asimismo, se ha observado que los terneros y el ganado lechero no pueden soportar grandes cantidades de moscas sin sufrir daños, de aquí que más de 50 moscas en vacas lecheras se consideran de importancia económica. Particularmente se considera que el UE para vacas Holstein es de 80-100 moscas por animal(44). Las vacas de carne pueden tolerar más de 200 moscas por animal, mientras que los toros pueden tolerar mayor cantidad de moscas(45). En términos generales, el UE para las infestaciones por H. irritans en bovinos es de ≥200 moscas por animal(16,42). En México, la estimación del UE se realiza generalmente mediante la observación visual y se han encontrado picos máximos en promedio de 300 y 120 moscas /animal en el sureste y centro del país respectivamente, ocurriendo estas infestaciones en la época de máxima precipitación pluvial(2).
Control químico y métodos de aplicación
Control químico
El tratamiento químico es el principal método de control de H. irritans. Las familias de insecticidas que se usan para el control de H. irritans en el ganado bovino son:
Organofosforados (OFs). Los insecticidas OFs son derivados del ácido fosfórico e interfieren con la función nerviosa a nivel sináptico inactivando la acetilcolinesterasa (AChE) que reacciona con los residuos de serina localizados en el sitio de catálisis de la AChE y al no hidrolizar a la acetilcolina, ésta se acumula de forma excesiva, generando incremento del estímulo y parálisis del insecto(46). Este mecanismo hace que los OFs sean también altamente tóxicos para los animales y el humano. Los OFs actúan contra ectoparásitos de animales tales como moscas, pulgas, piojos, ácaros y garrapatas y fueron los primeros en utilizarse en el control de H. irritans. Los compuestos OFs más utilizados son el diazinón y el ethión, que se utilizan generalmente en el control de poblaciones de H. irritans resistentes a los piretroides(33). Los aretes con diazinón al 21.4 % tuvieron una reducción del 87 % de H. irritans en bovinos en pastoreo de Tuxpan, Veracruz, hasta por 90 días(47).
Piretroides (Ps). Los Ps son derivados de las piretrinas, insecticidas naturales que se encuentran en las flores de plantas Chrysanthemum cinerariaefolium. Las piretrinas se clasifican en piretrinas TI y TII. Las piretrinas TI carecen del grupo α-ciano que se localiza en la posición del alcohol fenil-benzilo de las piretrinas TII. Las piretrinas naturales son sensibles a la luz solar, no así en los Ps sintéticos(48). Los sitios blanco de los Ps son los canales de sodio y cloro donde actúan inhibiendo la transmisión del impulso nervioso de los insectos, causando cambios en la permeabilidad de membrana(33). Los Ps TI cambian la conformación de los canales de sodio de las membranas neuronales, en respuesta al estímulo, mientras que los TII afectan los canales de cloro, incluyendo los dependientes del ácido gamma amino butírico (GABA) con una despolarización de las membranas y supresión del potencial de acción(48). Debido a que los insectos tienen un mayor número de canales de sodio sensible, a sus estructuras y temperatura corporal, los Ps son altamente tóxicos para ellos, mientras que en los mamíferos la toxicidad es relativamente baja(48).
Fenilpirazolonas. Son componentes químicos del tipo fenil pirazola, y su principal representante para el control de moscas es el fipronil. Estos plaguicidas actúan sobre los receptores del GABA provocando el bloqueo de los canales del cloro. Asimismo, bloquean dos tipos de activadores glutamato de los canales del cloro que se encuentran solamente en invertebrados. Posteriormente se produce una parálisis y finalmente la muerte de los artrópodos. La formulación por derrame dorsal a base de fipronil al 1% tiene una eficacia >80 % para el control de H. irritans hasta 21 días después del tratamiento(49).
Lactonas macrocíclicas (LM). Las LM se dividen en tres familias, a) las avermectinas, productos de la fermentación de Streptomyces avermitilis, ejemplo: ivermectina, doramectina y eprinomectina, b) las milbemicinas, derivadas de la fermentacion S. cyanogriseus, ejemplo: moxidectina y c) espinocinas derivado de Saccharopolyspora, ejemplo spinosad(50). Las LM interactúan de forma irreversible con el GABA y los receptores glutamato del canal de cloro incrementando la conductividad de membrana, ocasionando parálisis de insectos y ácaros(51). Dado que las LM son efectivas contra endo y ectoparásitos se conocen como endectocidas. La composición química de las avermectinas y milbemicinas no se altera a su paso por el tracto digestivo y son excretadas de manera intacta, por lo que afectan el desarrollo de las larvas en el estiércol de animales tratados. Sin embargo, su principal desventaja es su eliminación a través de la leche(33). En bovinos en pastoreo de Tuxpan, Veracruz, la ivermectina inyectable tuvo una eficacia en la reducción de moscas superior al 90 % hasta por 90 días(47).
Reguladores del crecimiento (RC). Los RC de insectos son substancias que actúan acelerando o inhibiendo los procesos fisiológicos esenciales del desarrollo normal de los insectos o de su progenie. Los RC no necesariamente son tóxicos, más bien causan anormalidades que comprometen la sobrevivencia de los insectos(2). Un ejemplo son las hormonas juveniles (HJ), análogas de la ecdisona, que son específicas de los insectos. En cada fase evolutiva, la cantidad de HJ se reduce y de esa manera el insecto se desarrolla hasta adulto. Cuando la HJ está presente de manera constante se previene la maduración del insecto(52). El metropeno y la ciromazina se incluyen en este grupo no tóxico para los mamíferos y por lo tanto se aplican como bolo o suplemento alimenticio.
Inhibidores del crecimiento (IC). Los IC de los insectos bloquean la polimerización de N-acetilglucosamina, evitando así la síntesis de quitina, componente esencial del exoesqueleto, de esta manera previenen la emergencia de H. irritans(53). En este grupo se incluyen las benzoil-fenil ureas como el diflubenzuron, el lufenuron y el triflumuron, de los cuales el diflubenzuron es el más utilizado contra H. irritans. Estos productos actúan contra huevos y larvas, no contra las fases adultas, por lo tanto, su administración es por vía oral, ya sea en forma de bolo o como suplemento en sales minerales y suplementos, aunque existen las formulaciones en aspersión o en polvo. En EUA y Brasil, el diflubenzuron ha mostrado una reducción del 90 al 99 % de H. irritans en 20 a 33 días del tratamiento(53). En México, se usa el diflubenzuron oral en bovinos (1 g/animal/día) con buenos resultados para el control de H. irritans.
Derivados del pirrol. Los pirroles halogenados son compuestos orgánicos aromáticos producidos por Streptomices, conocidos también como proinsecticidas, ya que son activados por oxigenasas como el citocromo p450 a metabolitos más tóxicos una vez dentro del insecto. El blanco del pirrol es la mitocondria, afectando la fosforilación oxidativa, rompiendo el gradiente de protones e impidiendo la producción de ATP(54). A este grupo pertenece el clorfenapir, primer insecticida utilizado al 30 % en aretes para el control de H. irritans, que fue ampliamente utilizado como alternativa para el tratamiento de H. irritans resistentes a los piretroides(33).
Repelentes. Algunos extractos y aceites esenciales de plantas han mostrado actividad repelente contra insectos y representan una alternativa para disminuir el uso de insecticidas convencionales en unidades de producción preocupadas por producir productos orgánicos, o contar con alternativas de control diferentes para mitigar la resistencia a insecticidas. Se trata de compuestos nitrogenados o alcaloides, fenólicos, inhibidores de proteínas y aceites esenciales(55). Una limitante de estos extractos o aceites esenciales es que tienen corto tiempo de repelencia, por ejemplo, los aceites esenciales del zacate de limón (Cymbopogon citratus), geranio (Geranium odoratissimum) y menta (Menta piperita) al 5 % en aceite de girasol mostraron repelencia contra H. irritans de 8 a 24 h(56).
Atrayentes. Los atrayentes son substancias volátiles y pueden ser detectados a largas distancias por otros insectos por llevar señales de alarma o reproductivos. Se trata de feromonas o mensajeros químicos que se encuentran en la cera cuticular de las hembras de H. irritans. La cera cuticular está compuesta de cadenas de 21 a 29 carbonos y son estimulantes de la cópula para el macho(2). Las feromonas sintéticas al aplicarse en trampas tratadas con insecticidas han sido usadas para atraer insectos, pero hasta el momento en H. irritans las trampas funcionan como un método de control físico y no químico(57).
Métodos de aplicación
Existen varios métodos de aplicación de insecticidas para controlar H. irritans. El método de elección depende de factores como el tipo de explotación, el sistema de producción (intensivo, extensivo, mixto), fin zootécnico del ganado (carne y leche, o doble propósito), manejo de excretas, infraestructura, instalaciones y el personal técnico a cargo de la aplicación de insecticidas(2). Los métodos más comunes de aplicación de insecticidas se describen a continuación(3,58,59).
Aretes impregnados con insecticida: Se trata de aretes de plástico con uno o más insecticidas insertados en la matriz del arete, que a medida que este se mueve, el insecticida se libera en pequeñas cantidades, y se distribuye a través del pelo del animal. Los aretes existentes contienen Ps, OF, LM y mezclas de Ps y OF. Todos los animales de un hato deben utilizar los aretes y éstos deben retirarse si no se observa efectividad.
Polvos: Se trata de sacos o bolsas con el insecticida que se libera a través de filtros cuando el ganado está en contacto con la bolsa. Para este tipo de control se requiere que las bolsas sean suspendidas cerca de las tomas de agua y debe asegurarse que el polvo caiga sobre los animales. Los polvos también se utilizan en el tratamiento de las excretas.
Derrame dorsal: Los insecticidas de derrame dorsal (pour-on), se aplican a lo largo de la línea de la espalda de los animales, a dosis basada en el peso del animal. Este es uno de los métodos más usados en México en el ganado bovino.
Aspersión: Los tratamientos por aspersión son efectivos en el control de las moscas, pero es importante cubrir por completo a los animales. La aspersión incrementa el manejo y estrés de los animales, lo que representa una desventaja en su utilización; sin embargo, es un método a considerar cuando la cantidad de animales no es muy grande. La aspersión es uno de los métodos más usado en México.
Larvicidas orales: Los larvicidas orales se aplican en el alimento o en bloques minerales o en suplementos alimenticios. Estos insecticidas pasan del tubo digestivo a las heces donde previenen que el desarrollo de las larvas a la fase adulta; sin embargo, si la cantidad de producto adecuada no es ingerida, las moscas pueden mantenerse por encima del UE. Una solución a ello son los bolos de liberación lenta, los cuales se alojan en el retículo y el producto se libera de manera continua. Por la vía oral se aplican las LMs, los RC y los IC.
Inyectables o sistémicos: Aunque la gran mayoría de insecticidas se emplea por vía tópica, la inyección intramuscular es un método de aplicación de las LM como la ivermectina, muy usual en ganado de carne para el control de garrapatas, moscas y nematodos gastrointestinales.
VetGun®: Es una forma de administración novedosa, en la que se utiliza una pistola diseñada para disparar una cápsula de gelatina (VetCap®), que contiene una dosis de insecticida. Esta cápsula es muy frágil y se rompe por el impacto generado con el contacto con la piel, liberando el insecticida que comienza a migrar a través del pelo y piel del animal. Mediante este sistema se pueden aplicar cápsulas de 5 a 10 m de distancia de los animales. Sin embargo, es posible que el insecticida no cubra los dos lados del animal adecuadamente. Este método de control no está disponible en México de forma comercial, pero es un novedoso método que podría usarse en el futuro cercano.
Bioinsecticidas. Los bioinsecticidas son extractos, o aceites esenciales obtenidos de plantas que pueden ser utilizados en el control de H. irritans. Existe evidencia de que ciertos extractos de plantas como del neem (Azadirachta indica) que contiene azadiractina cuando se administra oralmente a los bovinos a dosis de ≥0.03 mg por kg de peso vivo por día y el molido de semilla del neem administrado como suplemento de ≥10 mg de semilla por kg de peso vivo, inhibe el desarrollo de H. irritans en las heces(60). Otro compuesto botánico que ha demostrado buena eficacia contra H. irritans es el p-anisaldeido, extraído de varias plantas tales como Pimpinella anisum y Cuminum cyminum(61), así como los aceites esenciales de Carapa guianensis(62), Eucalyptus polybractea(63) y Pelargonium spp.(56).
En un estudio realizado en vacas lecheras infestadas naturalmente con H. irritans en México, se encontró una reducción de 9.5 a 68.0 % de infestación después de asperjar el extracto de hoja de Larrea tridentata al 20%(64). En México se requiere identificar moléculas bioactivas presentes extractos de plantas nativas de las diferentes regiones del país, así como las formulaciones de vehículos y vías de administración en bovinos. En México se requiere identificar moléculas bioactivas presentes extractos de plantas nativas de las diferentes regiones del país, así como las formulaciones de vehículos y vías de administración en bovinos.
Resistencia de H. irritans a los insecticidas
La resistencia es una respuesta genético-evolutiva de poblaciones de insectos expuestas al estrés continúo debido a la frecuente exposición a los insecticidas. En el campo se sospecha de resistencia cuando un producto que antes era útil para el control, ya no demuestra el mismo efecto, siempre y cuando se asegure que el insecticida se está usando bajo óptimas condiciones de aplicación(42). Debido a que el ciclo biológico de H. irritans es de pocos días, los tratamientos se realizan a intervalos cortos, lo que provoca que la frecuencia de individuos resistentes se incremente de manera progresiva y el insecticida eventualmente pierda su efectividad biológica(20).
En H. irritans existen varios mecanismos de resistencia que se manifiestan como cambios en el comportamiento del insecto para evadir la exposición al insecticida, la detoxificación por sobreexpresión de la enzima citocromo p450 y la insensibilidad en el sitio de acción debida a mutaciones en el canal de sodio(42). La resistencia de H. irritans a los Ps está asociada a la resistencia al derribe debido a una mutación en el canal de sodio, la cual previene o reduce la interacción con el canal de sodio, estas mutaciones se conocen como kdr o super kdr(65). La resistencia a los OFs se debe a mutaciones puntuales que producen cambios en la estructura, conformación y sitio de acción de la acetilcolina. Estos cambios se han encontrado en el sitio activo de la AChE de mosquitos y mosca doméstica resistentes a los OFs y se sabe que producen una disminución de la sensibilidad de la AChE(33).
El diagnóstico de resistencia a insecticidas más utilizado es el bioensayo donde las moscas recién capturadas son expuestas a papeles filtro impregnados con insecticida a concentración letal (CL) 0, 50 y 99, usando acetona como diluyente. El ensayo se realiza en tres réplicas y después de 1 hora se obtiene el porcentaje de mortalidad por cada concentración y se compara con el testigo, impregnado sólo con acetona(66).
En la detección molecular de resistencia a Ps se amplifica un fragmento del gen que codifica para el canal de sodio de un solo individuo y mediante el análisis de las secuencias obtenidas se determina si las mutaciones son del tipo kdr o super-kdr. En el caso de los OFs, una mutación puntual donde una glicina es substituida por una alanina en la posición 262 de la secuencia de aminoácidos de la AChE es determinada por PCR y puede ser usada para la detección de moscas resistentes en el campo(67).
La resistencia de H. irritans a los insecticidas se documentó desde los años 60s del siglo pasado en EUA y en los 80s a los Ps en poblaciones de H. irritans controladas mediante aretes con Ps en Florida, EUA. En México, el primer estudio sobre resistencia de H. irritans se realizó en el Golfo de México(68), encontrando una alta resistencia al fenvalerato y en menor proporción a los OFs. En Tamaulipas, México, se realizó un estudio para conocer la susceptibilidad a la cipermetrina y al diazinón y para detectar la presencia de genes kdr y super-kdr. Sólo en un rancho se encontró el gen super-kdr, mientras que la frecuencia de kdr fue del 43 al 78 % de las poblaciones de H. irritans estudiadas(66). En otro estudio en Tamaulipas, México, mediante ensayos con papel filtro se determinó que la resistencia a los Ps estaba distribuida en todo el estado y sólo en el sur se encontraron poblaciones resistentes tanto a Ps como al diazinón(69). Posteriormente en el norte de Veracruz y el centro de Nuevo León, se obtuvieron resultados similares, una alta resistencia a los Ps y leve al diazinón(70). En Tierra Caliente, Guerrero, se encontró resistencia tanto a los OFs como a los Ps en H. irritans en el 100 % de 30 ranchos analizados(71). Actualmente no se conoce en México la distribución de la resistencia de H. irritans a las principales familias de insecticidas, por lo que es necesario hacer un estudio nacional del monitoreo de resistencia en distintas regiones del país.
Control alternativo
Control físico. Para el control físico se usan las trampas para moscas adultas, que tienen la finalidad de atraer a las moscas en los vuelos que realizan en búsqueda del ganado. Se han diseñado trampas en forma de cilindro o de copas invertidas, recubiertas con material adherible, bolas negras y brillantes con luz violeta, o bien impregnadas de otros atrayentes como feromonas(72). Otro diseño es la trampa de paso, que consiste en un túnel oscuro por el que transitan los animales, lo que provoca que las moscas se separen del ganado y vuelen hacia las zonas con luz, congregándose en el techo del túnel donde mueren en 2 a 12 h. Las trampas túneles con un sistema eléctrico de aspiración para las moscas representan una alternativa de control atractiva, con el inconveniente de la instalación y costo de la electricidad(34). En México el uso de trampas es limitado y representa un área de oportunidad para su desarrollo y evaluación. Con el control físico de H. irritans se reduce el uso de insecticidas y con ello limita la selección de poblaciones resistentes a los insecticidas.
Control biológico. El uso de enemigos naturales para el control de las poblaciones de H. irritans ha sido ampliamente explorado tanto en ensayos in vitro como en infestaciones controladas y naturales(73-76). Dentro de los enemigos naturales se incluye a los parasitoides Pteromalidos que parasitan de pupas de moscas como los incluidos en los géneros Muscidifurax spp. y Spalangia spp.; las bacterias entomopatógenas como Bacillus thuringiensis, los nematodos entomopatógenos como los géneros Steinernema spp. y Heterorhabditis spp., los hongos entomopatógenos de los géneros Beauveria spp., Metarhizium spp. e Isaria spp. Asimismo, los escarabajos estercoleros de la familia Scarabaeidae juegan un papel importante en el biocontrol de H. irritans al degradar las heces y al incorporar las heces al suelo, lo que no permiten el desarrollo de la fase larvaria de la mosca(50). Estos agentes de biocontrol tienen un riesgo mínimo hacia los invertebrados que no son blanco específico, así como para aves y mamíferos, además de que con su uso se disminuye el uso de insecticidas y como consecuencia la resistencia a ellos(77,78).
Los parasitoides existen en el mercado mexicano e internacional, disponibles para utilizarse en las unidades de producción pecuaria, ya que atacan a cualquier mosca. Se distribuyen en bolsas de tela o recipientes de plástico que contienen pupas de mosca doméstica parasitadas por uno o dos géneros de microavispas (Muscidifurax y/o Spalangia). Los recipientes se reparten en potreros y corrales 48 horas antes de la emergencia de los parasitoides adultos, que se establecen fácilmente en lugares con un uso moderado de productos químicos. En México se han reportado diversas especies, siendo los más frecuentes Spalangia endius, S. nigroaenea y Muscidifurax raptor(73).
Las bacterias entomopatógenas como Bacillus thuringiensis han demostrado ser una herramienta útil para el control de estadios larvarios de H. irritans, al aplicarse directamente al estiércol; sin embargo, las experiencias sobre su uso en el campo son limitadas. Los nematodos entomopatógenos (Steinernema spp. y Heterorhabditis spp.), se presentan como una alternativa de biocontrol que requiere de mayor investigación para alcanzar su uso en el campo(74,76).
En México, se han evaluado in vitro diversos aislados de B. bassiana (Bb), M. anisopliae (Ma) e Isaria fumosorosea para el biocontrol de H. irritans(75). Un estudio desarrollado en condiciones de infestación controlada en el trópico seco, usando diferentes formulados asperjados en el ganado, se demostró que cinco cepas de M. anisopliae controlaron del 94 a 100 % de la infestación entre los días 12 y 13 post-tratamiento, mientras que tres cepas de I. fumosorosea disminuyeron la generación de fases inmaduras de 90 a 98 % en el día 13 post-tratamiento(79).
Se evaluó un formulado acuoso de la cepa mexicana Ma134 de M. anisopliae en ganado infestado naturalmente con H. irritans en ganado lechero mantenido en clima semiárido, alcanzando una efectividad en el control de la infestación de 68% en cuatro semanas de tratamiento(80). De manera similar, se evaluó la cepa Ma135, contra infestaciones naturales de Stomoxys calcitrans y H. irritans, en ganado lechero mantenido en un sistema de pastoreo y confinamiento en corral. Los resultados indicaron una eficacia a las seis semanas de 69 y 58 % para S. calcitrans y H. irritans, respectivamente(81). La principal desventaja de la aplicación de hongos entomopatógenos es que los conidios se desactivan con los rayos ultravioleta, por lo que su uso práctico requiere de formulaciones que les protejan de la luz solar y que las aplicaciones se realicen antes de la salida del sol, para mantener su efectividad.
Los escarabajos estercoleros de la familia Scarabaeidae, al degradan materia orgánica en las heces, compiten con H. irritans por espacio y materia orgánica. Durante su proceso de apareamiento, los escarabajos entierran el estiércol en el suelo, evitando de esta manera que las moscas se reproduzcan. En condiciones de laboratorio el escarabajo Aphodius lividus es capaz de reducir la emergencia de H. irritans de 98 a 100 %(82). En otro estudio realizado en Norteamérica se encontró que un número mayor a 40 adultos de Digitonthophagus gazella en el excremento de bovinos redujo la emergencia de H. irritans de 38 a 56 %(83).
Las LM, principalmente ivermectina y doramectina afectan la sobrevivencia de las poblaciones de escarabajos estercoleros(59,84). La moxidectina al 10% en bovinos reduce la capacidad reproductiva del escarabajo Onthophagus landolti(85). Por lo anterior, se recomienda utilizar tratamientos selectivos que generan una menor excreción de LM, y como consecuencia una menor afectación a las poblaciones de escarabajos estercoleros.
Control inmunológico. La necesidad de métodos de control más amigables con el medio ambiente y la salud pública, ha propiciado la investigación sobre la respuesta inmune del ganado a los antígenos de H. irritans como un camino hacia las vacunas contra la mosca de los cuernos, similar al enfoque utilizado con garrapatas. Se ha demostrado que un nivel de infestación de 200 moscas por animal produce una débil respuesta de anticuerpos a los antígenos de la saliva de las moscas, que aumenta cuando se retiran las moscas de los animales. Esto sugiere un efecto modulador de los antígenos en las glándulas salivales de H. irritans(86). En otro estudio se encontró una correlación entre la reducción de huevos y los niveles de anticuerpos contra H. irritans alimentadas con sangre de bovinos inmunizados con antígenos de intestino de moscas; sin embargo, la mortalidad de moscas alimentadas de animales inmunizados no fue significativa(87).
Aun cuando la vacunación de bovinos con proteínas recombinantes como la trombostasina, una proteína inhibidora de la coagulación, que fue identificada en las glándulas salivales de H. irritan(88) y la hematobina, una proteína inmunomoduladora de la saliva, demostraron una disminución en el consumo de sangre, retraso en el desarrollo de huevos y disminución del número de moscas adultas. La vacunación experimental con hematobina recombinante aumentó la respuesta de IgG anti-hematobina en el ganado y redujo el número de moscas en un 30 % en comparación con los controles(4). Hasta el momento, se han evaluado muy pocas proteínas recombinantes y aún no existe una vacuna recombinante contra H. irritans.
Los estudios de genómica funcional y la proteómica brindan la oportunidad de descubrir nuevos antígenos candidatos vacunales que posteriormente puedan ser expresados y producidos en proteínas recombinantes para ser usadas solas o combinadas en ensayos de vacunación y reto contra infestaciones por H. irritans en bovinos. En México, se identificaron candidatos para el desarrollo de vacunas contra H. irritans, mediante silenciamiento de genes usando la interferencia de ARN (iARN) en una geneteca de ADNc construida de tejidos abdominales de H. irritans parcialmente alimentadas. La iARN del grupo funcional de inhibidor de proteasas mostró una alta mortalidad de H. irritans y una reducción en la oviposición posterior a la iARN de vitelogenina, ferritina y ATPasa. También se obtuvo una alta mortalidad de moscas adultas, así como reducción de la oviposición posterior a la iARN de los componentes del proteosoma, respuesta inmune y 5’-NUC. Sin embargo, estos candidatos no han sido evaluados en ensayos de inmunización contra H. irritans e infestación(89).
Se ha realizado poca investigación sobre la identificación de candidatos para el desarrollo de vacunas contra H. irritans y, hasta el momento, los resultados son preliminares. Por lo tanto, el control inmunológico de la mosca de los cuernos no es una alternativa a corto plazo. La reciente secuenciación, ensamblaje y anotación del genoma de H. irritans(90) será útil para la identificación de nuevos antígenos candidatos para el desarrollo de vacunas.
Control cultural, táctico, estratégico y selectivo
El control cultural es el uso de buenas prácticas de manejo como son la remoción y disposición adecuada de las excretas frescas de los corrales y establos, lo que interrumpe el ciclo biológico de las moscas y previene el desarrollo de nuevas poblaciones(42).
El control táctico es la acción inmediata, provocada por la ocurrencia de niveles de infestación considerados dañinos. Para poder lograr esto se debe hacer un monitoreo cada 8-10 días de la población de moscas y se realiza el tratamiento cuando el nivel de infestación sobrepase el umbral económico(2).
El control selectivo consiste en el tratamiento de aquellos animales que presenten mayores infestaciones por moscas dentro del hato. Barros et al.(91) realizaron varios ensayos con diferentes insecticidas al 25% y 50% del hato, encontrando una reducción de moscas en todo el hato y disminución del costo del tratamiento; sin embargo, los animales tuvieron que ser tratados con mayor frecuencia cuando la infestación de moscas persistió.
El control estratégico se basa en el conocimiento de la epidemiología de la infestación de H. irritans en una región para limitar las épocas de mayor infestación y daño económico tratando de evitar los picos poblacionales. Estos tratamientos pueden ser implementados una vez superado el nivel pre-establecido de infestación de poblaciones de moscas evaluadas semanalmente(2).
Control integrado
El control integrado de parásitos (CIP) consiste en la asociación del medio ambiente y la dinámica de población de las especies de parásitos, utilizando una combinación de técnicas y métodos sustentables que sean compatibles y que mantengan niveles bajos de las poblaciones de parásitos que causan pérdidas económicas. El CIP generalmente se asocia a una drástica disminución de la frecuencia de tratamientos y por consiguiente una disminución en la presión de selección genética y de parásitos resistentes(1).
El uso de diferentes estrategias específicas para el control de H. irritans ha sido explorada a nivel mundial; sin embargo, la integración de las estrategias no ha sido estudiada, como ha ocurrido con otros parásitos como garrapatas(92). El reto principal que existe a nivel mundial y en especial en México es el uso eficiente de un programa integrado de moscas hematófagas, mediante la implementación de diferentes estrategias de control químico y no químico.
Conclusiones
Con base en la información presentada y discutida sobre la situación y perspectivas de estudio de H. irritans en la ganadería bovina de México se concluye que:
La mosca de los cuernos H. irritans es un ectoparásito obligado del ganado bovino, que se distribuye por todo México durante el año, con picos en verano o época de lluvias. Este parásito es responsable de importantes pérdidas económicas en los sistemas ganaderos, destacando la necesidad de estudiar su dinámica poblacional en diferentes regiones del país, para establecer estrategias de control efectivas y evitar picos poblacionales.
El tratamiento químico es el principal método de control de las infestaciones de H. irritans. Los insecticidas disponibles para el control de estas moscas son los OFs, Ps, LM, RC, IC, pirroles, así como productos repelentes y atrayentes que se usan mediante diversos métodos y vías de aplicación. El uso frecuente de insecticidas favorece la selección genética de poblaciones resistentes de H. irritans. En México, poblaciones de H. irritans resistentes OFs y Ps han sido reportados en Tamaulipas, Veracruz, Nuevo León, Guerrero y sureste de México de resistencia a los tratamientos químicos.
El control biológico mediante el uso de B. bassiana, M. anisopliae e I. fumosorosea ha demostrado ser una alterativa para el control de H. irritans en la ganadería bovina mexicana. Los escarabajos estercoleros regulan las poblaciones de H. irritans al impedir que las fases inmaduras de las moscas continúen su ciclo; sin embargo, el uso frecuente de LM para el control de endo y ectoparásitos afecta su desarrollo. Se requiere aplicar un uso racional de las LM en la ganadería bovina y de esta manera preservar estos reguladores naturales de las poblaciones de H. irritans.
Se requiere realizar estudios para la identificación y desarrollo de nuevos bioinsecticidas para el control de H. irritans en el ganado bovino.
El uso de diferentes estrategias de control integrado de H. irritans ha sido poco explorada a nivel mundial y en México.
Literatura citada:
