https://doi.org/10.22319/rmcp.v13i2.5832
Revisión bibliográfica
Estrés por calor en ganado lechero con énfasis en la producción de leche y los hábitos de consumo de alimento y agua. Revisión
Abelardo Correa-Calderón a
Leonel Avendaño-Reyes a
M. Ángeles López-Baca a
Ulises Macías-Cruz a*
a Universidad Autónoma de Baja California. Instituto de Ciencias Agrícolas, 21751, Valle de Mexicali, B.C. México.
* Autor de correspondencia: ulisesmacias1988@hotmail.com, umacias@uabc.edu.mx
El impacto negativo del estrés por calor (EC) en la ganadería lechera repercute en cuantiosas pérdidas económicas a nivel mundial, dado que reduce la producción de leche, la eficiencia reproductiva y la vida productiva de las vacas. Adicionalmente, el mejoramiento genético continuo resulta en vacas muy productivas, pero menos tolerantes al EC debido a que producen mayor calor metabólico. Esto en conjunto con el calentamiento global convertirá al EC en un reto difícil de controlar para la industria lechera. Como respuesta dependiente del grado de EC, el ganado lechero realiza una serie de ajustes fisiológicos, metabólicos y conductuales como mecanismos de termorregulación para disipar el exceso de calor corporal y reducir la producción endógena del mismo, todo dirigido a mantener la normotermia. Sin embargo, la secreción láctea y la fertilidad se reducen por efecto directo de la hipertermia e indirectamente por la reducción en el consumo de nutrientes dietéticos. Los consumos de alimento y de agua están asociados estrechamente con la reducción de la productividad en ganado lechero expuesto a EC. Cabe mencionar que el impacto del EC en la productividad del ganado lechero varía entre razas, siendo las razas Bos taurus menos tolerantes al EC, particularmente la raza Holstein. Actualmente, se investiga en la identificación de genes asociados con la termotolerancia, los cuales son empleados en programas de selección por marcadores genéticos para producir vacas altas productoras de leche en climas cálidos. Por lo tanto, el objetivo de esta revisión es hacer un análisis comprensivo de los efectos del EC sobre la producción de leche, activación de mecanismos de termorregulación y conducta de ingesta en ganado lechero.
Palabras clave: Ganado Holstein, Termorregulación, Hipertermia, Cambio climático, Conducta de ingesta.
Recibido: 27/10/2020
Aceptado: 12/07/2021
El estrés por calor (EC) se define como la suma de fuerzas ambientales externas que actúan sobre el animal causándole un aumento en la temperatura corporal, y la activación de ajustes fisiológicos y conductuales en primera instancia(1). En ganado lechero, estos ajustes representan mecanismos de adaptación activados para tratar de mantener en equilibrio la homeostasis corporal(2). Conforme el calentamiento global continúe incrementando, la prevalencia del EC en ganado lechero también aumentará en términos de frecuencia, duración y severidad(3,4). Por lo tanto, la mitigación de los efectos negativos del EC en la productividad de los hatos lecheros se ha convertido en un reto para la industria lechera a nivel mundial(5).
El problema del EC es mayor en áreas geográficas cálidas donde puede comenzar a finales de primavera y prolongarse más allá de la época de verano, por lo cual una baja fertilidad y producción de leche en las vacas puede proyectarse hasta la época de otoño(2,6). La magnitud del EC está determinada por los efectos combinados de la temperatura ambiental (Ta), humedad relativa (HR), radiación solar y velocidad del viento(1,2). En el caso particular del ganado lechero, se calcula el índice de temperatura–humedad (ITH) a partir de la Ta y la HR para determinar en primer instancia el grado de EC al que están expuestos los animales, y en segunda, para tomar decisiones de manejo en el hato durante la época caliente(1,7). Bouraoui et al(5) informaron que la producción de leche declinó en 21 % cuando el ITH aumentó de 68 a 78 unidades, o bien cuando la Ta superó los 27 °C, sin importar la edad o etapa de lactación.
Dado el impacto negativo del EC en la fertilidad y producción de leche, este tema ha sido motivo de una infinidad de investigaciones en los últimos 50 años, y actualmente se tiene conocimiento profundo de los mecanismos fisiológicos, metabólicos, endocrinos y moleculares que conducen a que el EC desencadene una reducción en la productividad del ganado. Sin embargo, varios de estos mecanismos fundamentan su inicio en dos factores claves, la reducción en el consumo de alimento y el aumento en el consumo de agua(8). De hecho, la producción de leche se correlaciona estrechamente con el consumo de materia seca (MS) y agua en vacas lecheras sujetas a EC crónico(9). Los primeros ajustes fisiológicos y de conducta de ingesta en el ganado lechero son considerados mecanismos homeostáticos(8), y tienen como principal fin incrementar las pérdidas de calor evaporativas (i.e., respiración y sudoración) y reducir la producción de calor endógeno con el propósito de mantener normotermia(10-12). Cabe mencionar que el genotipo juega un papel importante en definir el grado de impacto que tiene el EC en la producción del ganado lechero(13,14), siendo el ganado Holstein una de las razas más susceptibles, mientras que el ganado nativo puro de regiones cálidas o sus cruzas con razas introducidas son más tolerantes(15-18). Por lo tanto, el objetivo de la presente revisión es hacer un análisis comprensivo de los efectos del EC sobre la producción de leche, la activación de mecanismos de termorregulación y la conducta de ingesta en el ganado lechero.
Cambio climático y el futuro de la producción lechera
La producción de gases de efecto invernadero tales como vapor de agua, dióxido de carbono, metano y óxido nitroso principalmente, alteran la permeabilidad de la atmósfera permitiendo la entrada de los rayos solares mientras que evitan la salida del calor radiado por la superficie terrestre(19). Así, estos gases son los causantes del problema del calentamiento global y, por ende, del cambio climático, el cual tiende a incrementar la cantidad de superficie terrestre con climas cálidos, y en casos extremos, la desertificación(20). Estudios conducidos por climatólogos sugieren que la Ta podría incrementarse hasta en más de 2 °C para el año 2050(19).
En la actualidad, la rentabilidad de la industria lechera se encuentra amenazada por el cambio climático, ya que este fenómeno ha causado una disminución en la disponibilidad de recursos naturales, pastura y producción de grano y forrajes, al mismo tiempo que ha favorecido las pérdidas de ganado por siniestros naturales, así como la prevalencia de parásitos y enfermedades inesperadas(2,21). El aumento de áreas agroecológicas con Ta elevadas debido al cambio climático también ha sido una situación que ha llevado a mermar la producción de leche y a incrementar los costos de producción(8,22). De hecho, se espera que el EC incremente en severidad, duración y presencia en un futuro cercano(21). En Estado Unidos, las pérdidas económicas en la industria lechera por efecto del EC se estimaron en 897 millones de dólares(22), y recientemente proyectaron pérdidas económicas de 0.4 y 1.2 % para el 2050 y 2100, respectivamente, en la cuenca lechera ubicada al noroeste de ese país(21). En México, no existen reportes de las pérdidas que genera el EC en el ganado lechero, sin embargo, son evidentes si se toma de referencia la estacionalidad que hay en la producción de leche en cuencas ubicadas en climas cálidos(6,23). Cabe mencionar que la seguridad alimentaria del mundo se encuentra comprometida por el cambio climático; esto considerando que la demanda de carne y leche aumentará en 73 y 58 %, respectivamente, para el 2050 en relación con los niveles de producción del 2010(24).
Termorregulación y producción de calor metabólico en ganado lechero
Los bovinos como cualquier animal de granja son homeotermos, por lo que tienen la capacidad de mantener su temperatura corporal interna relativamente estable independientemente de las condiciones ambientales de su entorno(25). En ausencia de algún insulto térmico o presencia de fiebre, la temperatura corporal del ganado lechero se ubica entre 38.0 y 38.5 °C, y es el reflejo combinado del equilibrio entre las ganancias de calor ambiental y la producción de calor metabólico en un estado de homeostasis(1). De hecho, en un ambiente termoneutral y sin producción láctea, las vacas pueden mantener su normotermia por usar principalmente medios no evaporativos (conducción, convección y radiación), sin que esto implique un aumento en el costo energético asociado a la termorregulación(26). Si la temperatura rectal (TR) llega a ser ≥42 °C, se puede romper el equilibrio homeostático dentro del cuerpo, resultando en la muerte del animal(22).
Cabe señalar que la temperatura del cuerpo no es estable y exhibe un ritmo circadiano que varía en alrededor de 1°C entre la temperatura máxima y mínima registrada a través del día(1). La temperatura corporal alcanzará su pico máximo entre las 8 y 10 h después que la Ta alcanzó su máximo nivel, pero la vaca lechera es capaz de adaptarse a cambios de Ta y HR a través del año(27,28). Esto se debe a que la zona termoneutral del ganado lechero es muy amplia, fluctuando en el rango de −0.5 a 20.0 °C(1,2,10). Algunos estudios indican que la Ta crítica máxima en la cual el ganado Holstein puede mantener estable su temperatura corporal se sitúa entre 24 y 27° C(29).
Por otra parte, la síntesis de leche en la glándula mamaria de la vaca demanda una gran actividad metabólica, y consecuentemente, la producción de calor endógeno se eleva en proporción con el nivel de producción leche(25,27). Resulta imperante señalar que el mejoramiento genético del ganado lechero se ha enfocado en incrementar los parámetros de producción de leche, y esto ha convertido a la vaca en una máquina de producción de calor metabólico, lo que la hace muy susceptible a climas cálidos(17,30,31). Las vacas que producen entre 18 y 31 L/día de leche generan entre 28 y 48 % más calor metabólico que una vaca seca(32). Una vaca con un peso de 700 kg y con una producción de 60 kg/día de leche produce alrededor de 44,171 kcal de calor por día; esta misma vaca produce 25,782 kcal de calor por día al final de la lactancia (20 kg/día)(27). Las vacas que convierten el alimento a leche más eficientemente producen menos calor metabólico y tienen una menor temperatura de la superficie de la piel(33). Así que las vacas alta productoras de leche tendrán que hacer un mayor esfuerzo para regular su temperatura interna en condiciones termoneutrales, mientras que en condiciones de Ta alta serán menos tolerantes al EC.
Termorregulación fisiológica del ganado lechero en estrés por calor
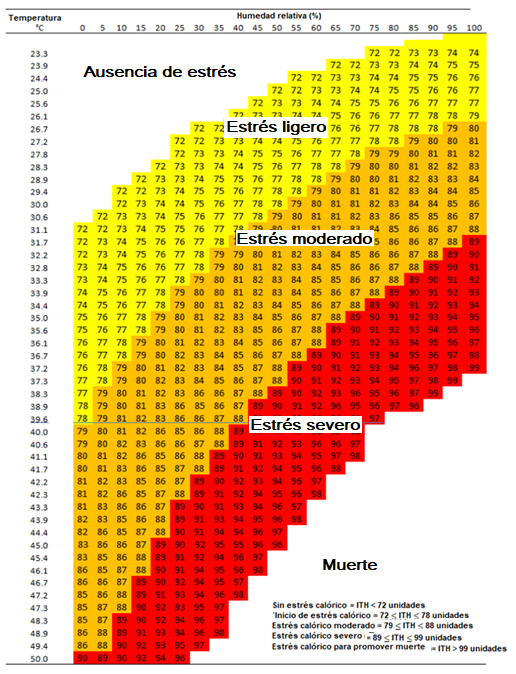
Las Ta altas en combinación con un elevado porcentaje de HR limitan aún más la habilidad de la vaca lechera para disipar el calor, y en consecuencia, experimentan EC y activación de mecanismos de termorregulación fisiológicos para disipar la carga de calor(2,5,26). Algunos índices climáticos han sido desarrollados para predecir el nivel de EC en ganado lechero, sin embargo, el ITH propuesto por Hahn(34) es el más usado a nivel mundial. Este índice combina los valores climáticos de Ta y HR en una ecuación (ITH = [0.81 × Ta] + [HR / 100] * [Ta – 14.4] + 46.4), y se considera que una vaca lechera comienza a presentar EC a las 72 unidades(1). De hecho, Armstrong(35) clasificó el EC en tres tipos de acuerdo al ITH: ligero (72 a 78 unidades), moderado (79 a 89 unidades) y severo (90 a 98 unidades) (Figura 1).
Aunque estudios recientes(11,36) reportaron que un ITH a partir de 68 sería el valor crítico en el cual la vaca lechera inicia a mostrar síntomas de EC. Otro estudio(37) encontró que las vacas lecheras de alta producción mostraron síntomas de EC cuando el promedio diario de ITH fue mayor a 68 unidades, o bien cuando el promedio mínimo de ITH fue superior a 65 unidades. El clima en que se desarrolle el ganado lechero también juega un papel importante para definir el inicio del EC basado en el ITH, ya que en condiciones tropicales se determinó el inicio del EC a un ITH>70 unidades(1) y en condiciones templadas los efectos negativos del EC comenzaron a un ITH>60(9). Esto sugiere que la presión de selección genética para mejorar la producción de leche se contrapone con la tolerancia al EC en ganado lechero(17).
Una respuesta inminente a la presencia de EC en ganado lechero es un aumento en la TR debido a una acumulación excesiva de calor corporal obtenido a partir del ambiente. Esto activa las pérdidas de calor corporal tanto por medios no evaporativos como evaporativos(6,10,26). Aunque dependiendo del tipo (agudo o crónico) y grado de EC (ligero, moderado o severo), será la secuencia de activación de los mecanismos de termorregulación fisiológicos(11). Conforme el EC es más severo y crónico, las pérdidas de calor corporal por medio no evaporativos disminuyen, mientras que las pérdidas evaporativas de calor por sudor y jadeo se incrementan en una forma considerable(8). Maia et al(26) observaron 85 % de pérdidas de calor corporal debido a la evaporación cutánea, y solamente 15 % a través del tracto respiratorio en vacas Holstein cuando fueron expuestas a Ta >30 ºC.
Figura 1: Clasificación el estrés calórico de acuerdo al índice de temperatura-humedad (ITH) en ganado lechero (adaptado basado en Armstrong, 1994)
La efectividad de las pérdidas de calor corporal por medios evaporativos se reduce cuando la HR ambiental es alta(10). Los resultados de Bonmanova et al.(7) demostraron que la HR fue el factor limitante del EC en climas húmedos, mientras que en climas cálidos secos fue la Ta. Adicionalmente, la activación de estos mecanismos termorregulatorios evaporativos implican un gasto extra de energía a diferencia de los mecanismos no evaporativos, por lo cual hay un aumento en el gasto energético de mantenimiento en las vacas lecheras estresadas por calor. Un incremento en la frecuencia respiratoria (FR) requiere de 7 a 25 % de gasto extra de energía debido al movimiento de los músculo del tracto respiratorio(38), y el proceso de vaporización de agua a través de la piel y tracto respiratorio demanda un gasto de energía de 2.43 J/ml de agua evaporada(1). Por lo anterior, la activación de pérdidas de calor por medios evaporativos representa para el animal una opción cuando las pérdidas no evaporativas son incapaces de liberar toda la carga de calor corporal bajo EC. En este sentido, la FR aumenta a partir de un ITH de 73 unidades, mientras que la TR a un ITH >80 unidades(39).
Por otra parte, las variaciones diurnas en las condiciones climáticas y los cambios en la conducta animal ayudan a mejorar la capacidad fisiológica de termorregulación del ganado lechero estresado por calor. Una Ta ≤21°C durante la noche, con duración entre 3 a 6 h, favorece la pérdida total del calor corporal acumulado en las horas del día, ya que el animal promueve una redistribución del flujo sanguíneo hacia la piel permitiendo una liberación eficiente de la carga de calor corporal por medios no evaporativos (radiación y convección)(40). Una velocidad del viento entre 1.8 y 2.8 m/s también beneficia las pérdidas de calor corporal a través de la superficie de la piel(28). Igualmente, las vacas adoptan una postura de pie para reducir el área corporal expuesta a los rayo solares, así como evitar la ganancia de calor a partir del contacto con el suelo(41). Ellas también bajo EC reducen la cantidad de alimento consumido por día y cambian sus hábitos de consumo hacia horas del día donde la Ta es más baja(8). Esto permite que su consumo de agua diario aumente(42).
Estrés por calor y consumo de alimento
La reducción en la producción de leche en ganado estresado por calor se asocia tradicionalmente con una reducción en el consumo de alimento(8) y su grado de disminución depende de la duración y severidad del EC prevaleciente, así como del grado de adaptación de la raza(14,18,25). Así, la Ta tiene una relación inversa con el consumo de MS bajo condiciones de EC crónico, mientras que, en condiciones de EC agudo, la ingesta de alimento se reduce rápidamente un día después de la exposición(43). De hecho, el consumo de alimento comienza a declinar a una Ta de 26 °C en vacas lactando(1), y su reducción puede alcanzar hasta el 40 % a Ta≥40 °C(44).
Dado que la presencia de EC se determina en base al cálculo del ITH en ganado lechero y este parámetro climático es más fácil de obtener que medir diariamente el consumo de alimento, algunos investigadores optaron por establecer la asociación entre ello y los resultados evidenciaron una disminución de 0.51 kg en el consumo de MS por cada unidad que incrementa el ITH dentro del rango de 72 a 84 unidades(8). Otro estudio encontró que la variación en el ITH mínimo tiene una mayor asociación con los cambios en el consumo de alimento que la variación del ITH máximo(45).
Aunado a la disminución del consumo de alimento, las vacas estresadas por calor también modifican los horarios de ingesta de la dieta con el objetivo de reducir su producción de calor endógeno en las horas del día más calientes. El calor metabólico asociado a la fermentación ruminal representa entre el 3 y 8 % del total de calor endógeno producido por el bovino, dependiendo del nivel de fibra dietética(12). La vaca lechera tiene diariamente un patrón natural de consumo de alimento, con consumos altos después de recibir alimento fresco, así como, por la tarde y al anochecer(46,47). Sin embargo, el ganado lechero bajo EC prefiere consumir alimento muy temprano por la mañana y al anochecer, ya que la digestión y producción de calor metabólico alcanza su pico entre 3 y 4 h después del consumo, justamente antes del periodo más caliente del día(32). Cabe mencionar que el EC en ganado lechero también impacta en la cinética ruminal del alimento consumido, ya que disminuye la motilidad del tracto gastrointestinal, cambia los patrones de fermentación y producción de ácidos grasos volátiles, e incrementa el tiempo de retención del alimento en rumen y su digestibilidad(38,47).
Algunos ganaderos con el propósito de mitigar la caída del consumo de MS y sostener la producción de leche durante las época caliente del año, modifican la composición química de la dieta para volverla más energética, lo cual implica incrementar el contenido de grano y reducir la cantidad de fibra dietética(12,39). Esta estrategia nutricional beneficia al ganado, ya que con un menor consumo de alimento puede llenar sus requerimientos nutricionales. Sin embargo, la modificación en ingredientes de la dieta puede tener consecuencias negativas si se excede en los ajustes; por ejemplo, una alta cantidad de urea en sangre debido a una elevada digestión de proteína en rumen incrementa la TR(48), o bien una dieta alta en concentrado puede coadyuvar a potencializar el problema de acidosis que genera el EC en las vacas(12).
Estrés por calor y consumo de agua
Una insuficiente disponibilidad de agua para beber tiene efectos adversos sobre la productividad y bienestar animal del ganado lechero bajo cualquier condición ambiental, aunque se vuelve más crítica a Ta elevadas(42). Independientemente del clima, las vacas lactando requieren grandes cantidades de agua, ya que tanto el cuerpo y la leche están compuestos por más del 80 % de agua(38,49). No obstante, estas vacas pueden variar en su nivel de consumo de agua de acuerdo a la cantidad de MS consumida en la dieta y kilogramos de leche producidos diariamente(50). Una restricción en el consumo de la misma puede reducir la producción de leche hasta en 26 % y bajo condiciones de EC esta reducción puede ser aún mayor(51).
La vaca lechera satisface sus requerimientos de agua por tres vías: consumo directo del agua, agua encontrada en los alimentos y agua derivada de la oxidación metabólica de tejido corporal(42). No obstante, la principal fuente de agua para el ganado proviene de la ingesta directa bajo cualquier condición climática (83 %)(49). El consumo de agua es afectado por factores como: consumo de MS, producción de leche, e ingesta de potasio, sodio y nitrógeno(52). Una vaca en condiciones termoneutrales puede consumir entre 14 y 171 kg/día de agua, con un promedio diario de 82 kg; basado en la producción de leche y consumo de sodio en la dieta, el consumo de agua se estima entre 4 y 4.5 L/kg de leche producido(42). Por otra parte, se encontró un consumo de 146 g de agua por cada gramo de sodio ingerido(52) en ausencia de un insulto termal. En vacas secas no estresadas por calor, el consumo de agua se reduce significativamente entre 15 y 61 L/día(53), pero notoriamente se incrementa en 27 % con el inicio de la secreción láctea en la lactancia temprana(54).
El ganado lechero puede experimentar una severa deshidratación cuando pierde el 12 % de su peso corporal debido a la falta de ingesta de agua, y puede resultar letal si la pérdida total de agua corporal es de aproximadamente 20 %(55). Dado que el EC aumenta considerablemente las pérdidas de agua en rumiantes, su ingesta a niveles adecuados se vuelve un punto crítico en las vacas lecheras(54). En zonas de climas cálidos, el consumo de agua puede incrementarse entre 10 y 20 % durante el verano, por lo que una vaca puede beber por arriba de 100 L/día, a pesar de ser una vaca clasificada como baja productora(32). Recientemente, un estudio reportó que el consumo de agua puede incrementarse hasta en 50 % conforme el ITH se aproxima a 80 unidades(56). Se estima que una vaca incrementa su ingesta de agua en 1.52 kg por cada grado celsius que aumenta la Ta(42), asimismo el tiempo dedicado diariamente a consumir agua aumenta de 0.26 a 0.5 h como el ITH cambia de 56.2 a 73.8 unidades(57) o la Ta de 15 a 33 °C(58). El mayor consumo de agua bajo condiciones de EC se atribuye a un aumento en el volumen de orina (25 %), evaporación del tracto respiratorio (jadeo) y pérdidas de calor por sudor(1), los cuales son mecanismos termorregulatorios que ayudan a disipar con mayor rapidez la carga excesiva de calor corporal en este tipo de ambientes(59).
Por otra parte, como previamente se señaló, los consumos de agua en el ganado lechero son muy altos bajo condiciones termoneutrales y aún más en EC, lo cual lleva a que esta especie cuente con conductas de ingesta de agua diferentes a otras. En condiciones termoneutrales, las vacas Holstein con producción media de leche consumen alrededor de ocho veces al día agua y en cada ocasión ingieren 12.9 L en promedio(60), mientras que las vacas Holstein altas productoras consumen con mayor frecuencia agua (12 a 16 veces) pero en menor cantidad en cada ocasión (5.4 a 6.0 L)(61). Estas vacas no estresadas prefieren consumir agua en horario diurno (97 % del total) que nocturno(60). Contrariamente, en condiciones de EC, la conducta de consumo de agua es muy variable entre los animales cuando la Ta excede los 30 °C(62). Probablemente, esto se deba a diferencias individuales en la tolerancia al calor, y por ende, al nivel de producción de leche. En general, las vacas estresadas por calor asisten al bebedero con mayor frecuencia que vacas no estresadas, pero ellas prefieren beber agua en horarios donde la radiación solar es baja o ausente(63). Esta conducta de bajo consumo de agua en horarios diurnos, particularmente en las horas más calientes del día (tres a cinco veces), obedece a que la vaca prefiere mantenerse inmóvil antes de cargarse de calor por la búsqueda de este nutriente. Por lo tanto, el diseño de construcción de los corrales o áreas de pastoreo se vuelve un factor crucial en la decisión de la vaca para ir a tomar agua bajo un clima caliente.
Los bebederos localizados a grandes distancias del área de comederos, especialmente cuando estos no poseen sombra, fomentan que las vacas bajo EC seleccionen entre movilizarse hacia al área sombreada o al bebedero después de comer(56). Por lo anterior, se recomienda que los bebederos sean distribuidos estratégicamente tanto en pastoreo y en corral para que las vacas no caminen más de 250 m(32); igualmente estos animales prefieren bebederos altos y largos(64). La limpieza y la temperatura del agua también son factores que definen en gran medida que las vacas expuestas a Ta alta consuman la cantidad de agua necesaria. El ganado lechero en general, al igual que cualquier otra especie, evita consumir agua mal oliente por lo que el lavado continuo de los bebederos es necesario bajo cualquier condición climática(60). Las vacas también prefieren ingerir agua un poco más caliente (≥24 ºC) que agua fría (≤10 ºC) o fresca (15 a 20 ºC), tanto en condiciones termoneutrales(65) como de EC(66). Cabe mencionar que esta preferencia de las vacas estresadas por calor por consumir agua caliente en lugar de fría resulta algo contradictorio, ya que se ha demostrado que el consumo de agua fría reduce tanto TR y FR(67). Dada la importancia de este nutriente, la medición del consumo de agua en ganado lechero estresado o no por calor es indispensable para mantener una producción de leche óptima, así como vacas saludables(42,50).
Impacto del estrés por calor en la producción de leche
La eficiencia óptima de producción en las vacas lecheras se da entre los 5 y 25 oC, por lo que se considera como la zona de confort de la vaca en lactación(1). La exposición de las vacas a Ta mayores al límite superior de esta zona de confort puede reducir la producción de leche entre 10 y 40 %(38). Aunque el impacto del EC en la producción de leche depende de la etapa de lactancia, la intensidad del calor y el potencial genético que tiene la vaca para sintetizar leche. Así, las vacas lecheras de alta producción en respuesta al EC presentan una mayor reducción en la síntesis de leche comparado con vacas medianas o bajas productoras(12,25). En respuesta al EC, las vacas de lactancia intermedia declinan su producción en 35%, mientras que las vacas de lactación temprana solamente en 14 %(25). Las vacas lecheras de alta producción tienden a presentar una mayor estructura ósea y un tracto gastrointestinal más grande, lo cual les permite la ingesta y digestión de más alimento, sin embargo, la producción de calor metabólico también aumenta y reduce la habilidad de la vaca para mantener su normotermia bajo condiciones de EC(1). De hecho, un incremento en la producción de leche de 35 a 45 kg/día reduce en 5 °C la Ta a la cual comienza el EC(68).
La producción de leche comienza a disminuir a una Ta de 27 °C sin importar la edad o etapa de lactación(1), y este efecto negativo se vuelve inminente entre 24 y 48 h posteriores al inicio de cualquier tipo de EC(43). Una investigación realizada con vacas Holstein Israelitas de 3era y 4ta lactancia, encontró una disminución de 0.38 kg/vaca/día en la producción de leche por cada grado centígrado que aumentó la Ta durante el verano(69). Como era de esperarse, al igual que la Ta, el ITH está asociado estrechamente en forma negativa (r= - 0.76) con la producción de leche(5), por lo que se estima que este parámetro productivo disminuye 0.41 kg/vaca/día por cada unidad de ITH que aumenta arriba de 68 unidades(40). No obstante, hay estudios que reportan menor caída en la producción de leche (0.2 kg/vaca/día) por cada unidad de ITH incrementada por arriba de 72 unidades(70). Probablemente, las variaciones en el nivel de producción de leche y en la Ta podrían explicar esta discrepancia entre estudios.
La caída en la producción de leche se da tanto por efectos directos como indirectos del EC. En los efectos directos se incluyen los ajustes en el metabolismo de energía y proteína para priorizar la disponibilidad de nutrientes en los procesos de termorregulación de la vaca en lugar de enviarlos a la glándula mamaria para la síntesis de leche(71). De igual manera, el EC incrementa la presencia de estrés oxidativo, lo cual a su vez altera la actividad metabólica y molecular de las células del tejido secretor mamario, y esto reduce la eficiencia celular de la síntesis de los diferentes componentes de la leche(30). De manera indirecta, el mecanismo está asociado con la reducción en el consumo de MS, ya que los requerimientos de energía de mantenimiento incrementan debido a la activación de los mecanismos de termorregulación fisiológicos en un escenario donde la disponibilidad de nutrientes corporales son reducidos como consecuencia del bajo consumo de alimento(3,38). Una reducción en la eficiencia de utilización de la energía para producción de leche entre 30 y 50 % ha sido reportada para vacas lecheras en climas cálidos(1).
El EC no solo afecta negativamente la producción de leche, sino también a su composición(56).Una reducción en el contenido de grasa y proteína combinado con un aumento en el conteo de células somáticas (CCS), son hallazgos comúnmente asociados al EC(5). Los resultados de estudio donde compararon condiciones termoneutrales de primavera versus EC severo de verano, mostraron que el contenido de grasa (3.58 %) y proteína (2.96 %) en leche fue menor en verano por 30 y 23 %, respectivamente(5). En ese mismo estudio detectaron que el CCS aumentó en 110 % debido al EC de verano. Otro estudio reportó que las vacas Holstein bajo EC de moderado a severo redujeron en 40 % el contenido de grasa en leche, mientras que el contenido de proteína se disminuyó en 17 %(1). Por su parte, Bernabucci et al(72) observaron que el porcentaje de caseína en leche fue ligeramente menor en verano que en invierno (2.18 vs 2.58 %). La reducción en el porcentaje de caseína puede ser la explicación de la baja producción de queso en esa época(73).
Respuestas adaptativas de razas lecheras al estrés por calor
El ganado Holstein es el genotipo por excelencia para la industria lechera mundial, sin embargo, tiene poca resistencia al EC. Los programas de mejoramiento genético para esta raza han sido enfocados en seleccionar rasgos asociados con la mejora en la producción y calidad de la leche, los cuales tienen un efecto antagónico con su habilidad para tolerar Ta altas(28). Así, el índice de heredabilidad de la producción de leche en vacas Holstein puede disminuir por efecto del EC, y en consecuencia, la respuesta de selección basado en este rasgo puede no ser la esperada en climas cálidos(17). A pesar de esto, el ganado Holstein se continúa usando en regiones cálidas porque mantiene una producción promedio de leche diaria mayor que al de otros genotipos adaptados a la región, principalmente en sistemas de producción tecnificados y semi-tecnificados(13,18).
Si bien el ganado Holstein ha mostrado mantener una mayor producción leche en comparación con razas como Simmental (20.4 vs 28.1 L)(13) y Jersey (26.6 vs 34.2 L)(16) en condiciones de EC tropical, su leche se caracteriza por tener CCS alto y un porcentaje de grasa y proteína bajo. Adicionalmente, la caída en la producción de leche debido a EC es más pronunciada en vacas Holstein, siendo un síntoma inequívoco de menor tolerancia a Ta elevadas(13,18). Por lo tanto, el uso de la raza Holstein en climas cálidos, particularmente en regiones tropicales y subtropicales, no puede ser justificado basado en el nivel de producción de leche, dado que las condiciones de EC reducen la calidad de este producto y compromete su estado de salud y bienestar.
Para producción de leche en climas tropicales se recomienda el aprovechamiento de los recursos zoogenéticos de la región incluyendo razas bovinas autóctonas e introducidas con gran adaptación al clima cálido-húmedo(31,74). Algunos estudios también sugieren los cruzamiento entre razas Holstein y cebú para generar ganado con un genotipo lechero tolerante a climas cálidos(10,15). Así, las razas lecheras adaptadas a EC se muestran más tolerantes a estas condiciones ambientales porque fenotípicamente cuentan con pelo más delgado y menos denso, así como mayor densidad de glándulas sudoríparas en la piel, lo que les permite liberar la carga de calor corporal con mayor facilidad por convección y radiación(14,75). Adicionalmente, este tipo de ganado bajo EC mantiene relativamente su consumo de alimento y actividad de la glándula tiroides, lo cual explica la menor variación en la producción de leche comparado con ganado no adaptado(75). En el trópico Mexicano, las principales razas usadas para producción de leche son Pardo Suizo, Jersey, Holstein, Gyr y Brahman, así como sus cruzas; no obstante, otros factores de manejo y alimentación junto con las condiciones climáticas conducen a que presenten una eficiencia reproductiva y productiva baja(76).
Por otra parte, en ganado adaptado a EC, sus células activan el factores de transcripción del choque térmico para coordinar los mecanismos protectores en respuesta al insulto térmico, incluyendo la expresión de genes, activación de proteínas del choque térmico (HSP por sus siglas en inglés) y metabolismo de carbohidratos(77). En ganado tolerante (Bos indicus) comparado con el no tolerante al EC (Bos taurus), las células presentan mayor expresión de HSP (principalmente HSP de 70 kDa(78) y 90 kDa(79)) en respuesta al EC, lo que mejora su viabilidad y actividad inmune(80). No obstante, se ha demostrado que las células mononucleares de ganado lechero pueden presentar una sobreexpresión de HSP-72 en respuesta a hipertermia, siendo contraproducente para su respuesta inmune y tolerancia al calor(81). Esto último se evidenció en un estudio donde células mononucleares de ganado Holstein y Pardo Suizo se cultivaron in vitro a 39 (normal), 40, 41, 42 y 43 ºC, y los resultados mostraron que las condiciones de hipertermia causaron una excesiva producción de ARNm de HSP-72 y baja síntesis de ADN en ganado Pardo Suizo en relación a lo observado en ganado Holstein(81). Contrario a lo esperado, estos hallazgos sugieren que el ganado Holstein resiste más las condiciones de hipertermia que el ganado Pardo Suizo; aunque este último puede mantener adecuada eficiencia productiva bajo condiciones de EC mientras no sea comprometida su capacidad de termorregulación y cause un aumento en la temperatura interna mayor a 40 ºC(81).
Marcadores genéticos asociados con termotolerancia en ganado lechero
Una gran cantidad de estudios se han enfocado en identificar la expresión de genes asociados con la termotolerancia en ganado lechero adaptado al EC, y dicha información está siendo utilizada para establecer programas de selección genética asistida por marcadores(82). Igualmente, se trabaja en la identificación de machos que expresen genes de termotolerancia, los cuales son seleccionados como sementales con el objetivo que transmitan estas características a su descendencia(28). Especial atención se está poniendo en buscar estos genes en ganado lechero que presente una producción de leche alta como el interés de la industria láctea es tener genotipos tolerantes al EC que produzcan altos volúmenes de leche de calidad.
En México, poco se ha estudiado en relación a la identificación de genes ligados a la termotolerancia del ganado lechero. Hernández-Corderos et al(83) identificaron siete genes (AVPR1A, Furin, IGFBP5, IGFBP6, PMCH, PRLR y STAT5B) asociados con la producción de leche de ganado Holstein estresado por calor en el Valle del Yaquí, Sonora, México; específicamente un polimorfismo de nucleótico único (SNP por sus siglas en inglés) dentro de cada gen que se relacionan con la vía de prolactina y somatotropina. Otro estudio(84) reportó la identificación de seis genes asociados con la producción de leche y su contenido de grasa y proteína (SFXN1, LOC781028, ANKRD31, LOC100296562, LOC107131388 y WDR41) en un análisis de asociación genómica amplia conducido en ganado Holstein criado en la región desértica de Mexicali, Baja California, México. Se requiere seguir haciendo más estudios del ganado Holstein Mexicano localizado en climas cálidos para buscar otros genes asociados directamente con rasgos de termoregulación que, combinados con esos ya identificados para producción de leche, sirvan como marcadores genéticos para identificar ganado tolerante al EC.
En Estados Unidos, una investigación reciente(82) identificó tres regiones genómicas (BTA5, BTA 14 y BTA15) que explican en gran parte la variación en la producción de leche de ganado Holstein estresado por calor. En esas regiones se identificaron 10 genes relacionados con la termotolerancia, ya que intervienen en diferentes procesos celulares en respuesta al EC, tales como: activación de las HSP (PEX16, HSF1, EEF1D y VPS28), reducción del estrés oxidativo (CDKN1B y DUSP16), modulación del proceso de apoptosis (MAPK81P1, CREB3L1), mantenimiento del ADN (TONSL) y termotolerancia (CRY2). Otras regiones genómicas asociadas con la presencia de genes de termotolerancia en el ganado Holstein estadounidense son BTA-24(85) y BTA-26(86), las cuales se relacionan con TR y producción de leche, respectivamente. En el caso de China, también hay evidencias de que su ganado Holstein es portador de SNP asociados a genes de termotolerancia como ATP1A1(87), HSP90AA1(88), HSF1, HSP70A1A y HSPB1(28).
En otras razas de ganado lechero también se han identificado algunos genes de termotolerancia al EC. En ganado Senepol se tiene identificado en gen haplotipo SLICK, el cual se ha introducido en raza Holstein con resultados positivos en su tolerancia al EC sin comprometer la producción de leche(89). En ganado lechero de la raza indígena Sahiwal, el cual es nativo de la India, se encuentran los genes HSP90AB1y HSPB8(90). Cabe mencionar que las razas lecheras del trópico mexicano al parecer no han sido tipificadas para detectar genes asociados con termotolerancia a Ta altas, por lo cual es un tema de investigación que podría desarrollarse en el futuro.
Conclusiones e implicaciones
El EC es un factor clave que condiciona el nivel de producción de leche en el ganado, siendo más notorio en razas sensibles a Ta altas. El ganado lechero comienza a experimentar EC cuando las condiciones ambientales promueven un ITH de 72 unidades, aunque razas altas productoras de leche pueden iniciar desde las 68 unidades. Frente al ambiente de EC, este ganado hace ajustes fisiológicos y metabólicos para disminuir la producción endógena de calor al mismo tiempo que libera el exceso de carga de calor corporal, sin embargo, la activación de los mecanismos de termorregulación disminuyen la capacidad de síntesis y secreción láctea de la vaca. La disminución en la producción de leche está asociada cuando menos en un 40 % con un bajo consumo de MS, y el resto con efectos directos en el metabolismo general y termo-resistencia celular. La alta presión de selección para rasgos asociados a la producción de leche ha llevado en la actualidad a tener ganado menos tolerante al EC, por lo que es necesario modificar dicha estrategia de mejoramiento genético tomando en cuenta la presencia de genes de termotolerancia. En este sentido, los trabajos de investigación de EC en ganado lechero deben enfocarse mayormente en buscar estrategias de mitigación del mismo, las cuales ayuden a reducir el costo energético debido a la activación de los mecanismos de termorregulación, así como aumentar el consumo diario de alimento.
Literatura citada: