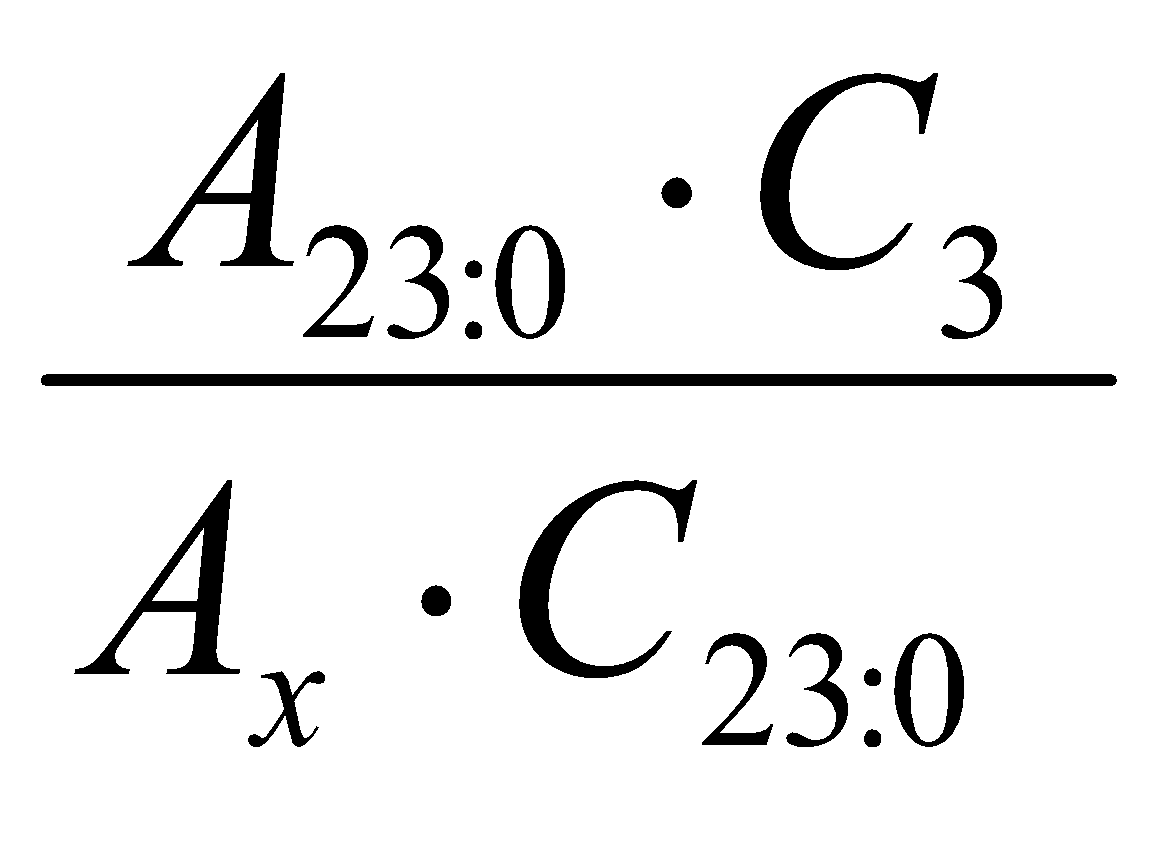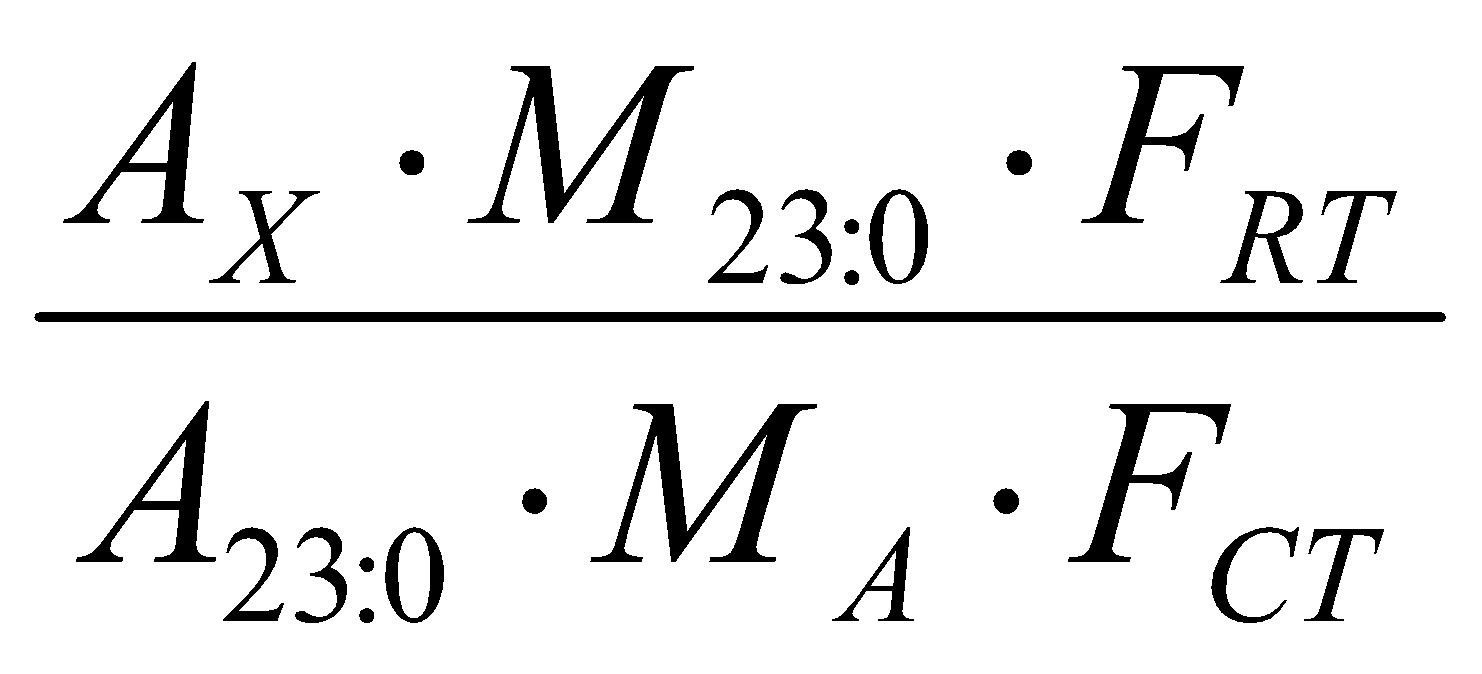https://doi.org/10.22319/rmcp.v12i1.4764
Article
Milk fatty acid profile of crossbred Holstein x Zebu cows fed on cake licuri
Antonio Ferraz Porto Junior a*
Fabiano Ferreira da Silva b
Robério Rodrigues Silva b
Dicastro Dias de Souza c
Edvaldo Nascimento Costa c
Evely Giovanna Leite Costa c
Bismarck Moreira Santiago c
Geónes da Silva Gonçalves c
a University of Southwest Bahia State - UESB. D.sc. Nutrition and Ruminant Production. INOVAPEC, Itapetinga, BA, 45700-000, Brazil.
b University of Southwest Bahia State - UESB, Department of Rural and Animal Technology, Itapetinga, BA, Brazil.
c University of Southwest Bahia State – UESB. Nutrition and Ruminant Production. UESB, Itapetinga, BA, Brazil.
*Corresponding autor: ferrazporto@hotmail.com
Abstract:
The objective was to evaluate the inclusion level of licuri cake (LC) in the diet of confined cows on fatty acid profile and milk cholesterol. Four cows with a blood level of ½ to ¾ Holstein x Zebu blood were used, distributed in a 4 x 4 Latin square, where the inclusion levels of the cake in the total diet were 0.0, 5.5, 11.0 and 16.5%, replacing soybean meal in the diet. There was a linear decreasing effect for the fatty acids Lauric, Elaidic, Gamma-Linolenic and conjugated linoleic acid isomers (C18:2cis9trans11; C18:2trans10cis12) from the inclusion levels of LC. The inclusion of licuri cake negatively influenced the concentrations of polyunsaturated fatty acids and conjugated linoleic acid isomers, in addition to the fatty acids of the Omega-6 series, which is not interesting from the human nutrition point of view.
Key words: Conjugated linoleic acid, Biohydrogenation, By-product, Cromatography.
Received: 08/02/2018
Accepted: 03/12/2019
Introduction
The licuri fruit (Syagrus coronata) (Martius) Beccari, is a typical palm tree of arid regions of the Caatinga biome. The cake is the main product after oil extraction(1), it can be used as an alternative source to lower production costs at a certain time of the year. However, the reduction in food costs depends of many factors, among others, proximity between ownership, by-product availability, nutritional characteristics and freight cost. Traditional ingredients used, such as corn, soybean, cotton, and biodiesel agribusiness by-products have been used as alternative sources.
The composition of the diet is the main factor influencing the fatty acid (FA) composition of meat and milk, since the fatty acids that reaches the duodenum are at least partly of food origin, as well as microbial bio hydrogenation of the diet dietary lipid rumen(2).
In order to meet the demand of an increasingly demanding market for the consumption of certain saturated fats, due to their negative effects on human health, the manipulation of milk constituents, especially in relation to fat has been increasingly studied(3), because they have some fatty acids, precursors of blood cholesterol (LDL) that are linked to cardiovascular problems. Thus, there is a real increase in the demand for healthy foods that has low levels of saturated fat and preferably those that are beneficial to human health.
However, human intake of essential FA and conjugated linoleic acid (CLA) decreased, reflecting their low concentrations in ruminant milk as well as the consumption of low fat dairy products(4). Therefore, the objective of this study was to evaluate the levels of inclusion of licuri cake in diets of confined cows on the fatty acid and cholesterol profile of milk.
Material and methods
Animals and treatments
The experiment was conducted on Fazenda Valeu Boi, located in the municipality of Encruzilhada – Bahia, between May 03 and August 22, 2016, 2016, and approved by the Ethics Committee on the Use of Animals (ECUA) under protocol nº. 104/2015 of April 15, 2015.
Four crossbred Holstein x Zebu cows (blood level between ½ and ¾ H x Z blood) were used, at the third or fourth lactation order, with average milk production between 4,500 and 6,000 kg in the previous lactation, adjusted for 300 d of lactation, with a mean body weight of 548 ± 17 kg. Cows were also selected according to lactation days, between 80 and 120 d at the beginning of the experimental period. They were arranged in a 4 x 4 Latin square, consisting of four experimental periods, with 21 d each, for which the first 16 d corresponded for adaptation and the last 5 d for data collection.
The licuri cake was purchased from Lipe Indústria de Sabão e Velas Ltda, Guanambi - Bahia. Inclusion levels of the by-product in the total diet were 0.0, 5.5, 11.0 and 16.5 %, which corresponded to the replacement of 0.0, 25.0, 50.0 and 75.0 % of the crude protein of the total diet.
The diets were formulated in an attempt to be isoenergetic and isoproteic, in order to contain sufficient nutrients for maintenance, body weight gain of 0.15 kg d-1 and production of 25 kg milk d-1, adjusted to 3.5 % of fat, according to the requirements table(5), and based on the chemical-bromatological composition data of sugarcane, corn, soybean meal and LC, previously analyzed before the beginning of the experimental period.
The bulk source used was sugarcane (Saccharum officinarum), variety RB 72-454, treated with 1% urea-ammonium sulfate mixture (9:1), on a fresh matter basis. Table 1 shows the proportions of the ingredients in the concentrates and the ratio bulk source:concentrate, on a dry matter basis.
Table 1: Proportions of ingredients based on dry matter
Ingredients | Levels of licuri cake (%DM) | ||||
0.0 | 5.5 | 11.0 | 16.5 | ||
Sugarcane | 49.9 | 49.8 | 50.2 | 50.1 | |
Ground corn | 35.4 | 32.7 | 29.7 | 27.0 | |
Soybean meal | 12.9 | 10.4 | 7.8 | 5.3 | |
Licuri cake | 0.0 | 5.3 | 10.6 | 15.9 | |
Minerals salt1 | 1.0 | 1.0 | 1.0 | 1.0 | |
Limestone | 0.6 | 0.6 | 0.5 | 0.5 | |
Dicalcium phosphate | 0.2 | 0.2 | 0.2 | 0.2 | |
Chemical bromatological composition | |||||
Dry matter | 94.2 | 91.7 | 91.1 | 91.7 | |
Crude protein | 23.3 | 21.1 | 20.7 | 20.2 | |
Ether extract | 5.81 | 6.2 | 5.9 | 6.6 | |
Neutral detergent fiber2 | 12.4 | 16.2 | 18.4 | 22.9 | |
Acid detergent fiber | 9.3 | 13.4 | 17.5 | 22.8 | |
Non-fibrous carbohydrates | 53.1 | 51.4 | 50.1 | 45.4 | |
Mineral matter | 5.4 | 6.2 | 6.1 | 6.0 | |
Lignin | 1.5 | 4.2 | 6.8 | 9.1 | |
*NDIN3 | 15.7 | 25.5 | 34.9 | 34.9 | |
*ADIN4 | 15.6 | 18.1 | 19.4 | 25.7 | |
NDFi5 | 1.1 | 5.4 | 9.9 | 13.9 | |
1Composition: Calcium 200 g, Cobalt 200 mg, Copper 1.650 mg, Sulfur 12 g, Iron 560 mg, Fluorine (max) 1.000 g, Phosphorus 100 g, Iodine 195 mg, Magnesium 15 g, Manganese 1.960 mg, Nickel 40 mg, Selenium 32 mg, Sodium 68 g, Zinc 6.285 mg, 2Corrected for ash and protein; 3Neutral detergent insoluble nitrogen; 4Acid detergent insoluble nitrogen and 5Indigestible neutral detergent fiber. *Values in percentage of dry matter of the total nitrogen.
The animals were allocated in covered individual 16m2 bays, with troughs and drinking fountains. The diets were provided to the animals in two daily fractions in the form of a complete mixture, always at the same time, at 0700 and 1400 h at will, in order to allow 5% of leftovers. In each experimental period, bulk source and supplements were collected to assess the fatty acid profile (Table 2). Sample lipid extraction of the samples was based on the procedure proposed(6)
Table 2: Lipid profile of sugarcane and concentrates consumed
Licuri cake levels (%DM) | |||||
Fatty acids1 | Fatty acid concentration2 (mg g-1) | ||||
Sugarcane | 0.0 | 5.5 | 11.0 | 16.5 | |
C4:0 | 104.8 | nd* | 0.0 | 0.0 | 0.1 |
C6:0 | nd* | nd* | 0.02 | 0.05 | 0.1 |
C8:0 | nd * | nd* | 0.1 | 0.3 | 0.4 |
C10:0 | nd* | nd* | 2.8 | 5.6 | 8.5 |
C12:0 | nd* | nd* | 1.7 | 3.4 | 5.1 |
C13:0 | 2.8 | nd* | nd* | nd* | nd* |
C14:0 | 11.0 | nd* | 10.2 | 20.3 | 30.7 |
C16:0 | 10.3 | 14.3 | 15.6 | 16.6 | 18.5 |
C18:0 | 7.1 | nd* | 1.2 | 2.5 | 3.7 |
C18:1n9t | nd* | 3.6 | 4.0 | 4.5 | 5.1 |
C18:1n9c | 14.4 | nd* | 2.0 | 3.9 | 5.9 |
C18:2n6 | nd* | 35.1 | 32.2 | 28.8 | 27.8 |
C20:1 | nd* | 0.8 | 0.7 | 0.7 | 0.7 |
C18:3n6 | nd* | 46.7 | 42.6 | 37.9 | 35.9 |
C21:0 | nd* | 2,3 | 1,9 | 1,6 | 1,3 |
C20:3n6 | nd* | 0.2 | 0.2 | 0.1 | 0.1 |
C20:3n3 | 0,6 | nd* | nd* | nd* | nd* |
C24:0 | nd* | 0.2 | 0.2 | 0.2 | 0.2 |
Totals fatty acids (mg g-1) | |||||
SFA3 | 135.9 | 16.8 | 33.8 | 50.6 | 68.6 |
MUFA4 | 14.4 | 4.3 | 6.7 | 9.1 | 11.7 |
PUFA5 | 0.6 | 81.8 | 74.9 | 66.8 | 63.9 |
1Usual nomenclature expressed in mg g-1 fat, Butyric (C4:0), Caproic (C6:0), Caprylic (C8:0), Capric (C10:0), Lauric (C12:0), Tridecanoic (C13:0), Myristic (C14:0), Palmitic (C16:0), Stearic (C18:0), Elaidic (C18:1n-9t), Oleic (C18:1n-9c), Gamma-linoleic (C18:2n-6), Eicosenoic (C20:1), Gamma-Linolenic (C18:3n6), Heneicosylic (C21:0), Dihomo-gamma-linolenic (C20:3n6), Eicosatrienoic (C20:3n3), Lignoceric (C24:0), 3saturated fatty acids, 4monounsaturated fatty acids, 5polyunsaturated fatty acids and *Not detect.
Analysis of fatty acids
Analyses were performed in the Laboratory of Chemical Separation Methods (LABMESQ), at the State University of Southwest Bahia (UESB). For the extraction of total lipids from fresh milk, 50 mL of each thawed sample were centrifuged at 12,000 rpm for 30 min at 4ºC, in a high-speed microcentrifuge (Himac CF-16RX II). The solid layer formed in the upper part was collected and stored in eppendorfs for further analysis, following the methodology proposed(7).
The lipids extracted from fresh milk were submitted to the preparation of fatty acid methyl esters, according to the procedure described(8), with modifications described(9). Fatty acid esters were analyzed using a gas chromatograph, (GC-2010 Plus Shimadzu), equipped with a Flame Ionization Detector (FID) and a Rt-2560 fused silica capillary column (100 m, 0.25 mm d.i). Gas flow (White Martins) was 40 mL min-1 for the carrier gas (H2); 30 mL min-1 for the auxiliary gas (N2) and 4,000 mL min-1 for the synthetic air flame. The sample split ratio was 90:10. The operating parameters were set after verification of the best resolution conditions. Injector and detector temperatures were 225 °C and 260 °C, respectively. The column temperature was programmed at 140 °C for 5 min, followed by of 3 °C min-1 ramp to reach 245 °C for 20 min. The total analysis time was 60 min. Injections were performed in duplicate and injection volume was 0.7 μL. The peak areas of fatty acid methyl esters were determined using the LCSolution® software.
Identification of methyl esters
Methyl esters were tentatively identified by the retention time, comparing the standard containing 37 fatty acid methyl esters (189-19 Sigma, USA) and the retention times of fatty acid methyl ester standards containing the c9t11 and t10c12 linoleic acid isomers (O-5632 Sigma, USA)(9).
In order to evaluate the response of the flame ionization detector, a mixture solution consisting of standards (Sigma) of fatty acid methyl esters was used at a known concentration and was calculated by the equation according to the method proposed by Ackman(10). These factors were obtained from the mean of four replicates:
FR =
Where: FR= Response factor in relation to methyl tricosanoate; A23:0= Area of methyl tricosanoate; C3= Concentration of fatty acid methyl esters; Ax= Area of fatty acid methyl esters and C23:0= Concentration of methyl tricosanoate.
For fresh milk samples, fatty acids were quantified in mg g-1 total lipids, using the internal standard methyl tricosanoate (23:0) (Sigma, USA). After weighing the lipids (~ 150 mg) for transesterification, 1000 μL of the internal standard solution at a known concentration (1.00 g mL-1), were added to all samples with the aid of a micropipette. The concentration of fatty acids in the samples was calculated according(11).
C (mg g-1) =
Where: AX= Area of fatty acid methyl esters; A23:0= Area of the internal standard; M23:0= Mass of the internal standard added to the sample (in milligrams); MA= Sample mass (in grams); FRT= Theoretical response factor of fatty acid methyl esters and FCT= Conversion factor to express the results of fatty acids, in mg, per g of total lipids (LT).
Extraction and identification of milk cholesterol
Extraction, detection, identification and quantification of cholesterol were performed, following the methodology of Bauer, et al(12). A C18 (250 mm x 4.6mm x 5 µm) analytical column was used. The mobile phase consisted of acetonitrile:isopropanol (95:5), at a flow rate of 2 mL min; the analysis time was 28 min. Chromatograms were run at 202 nm using an HPLC apparatus (Shimadzu), equipped with a degasser (DGU – 20 A5R) and two pumps (LC-20 AR), with a UV-Visible detector (SPD – 20 A). Cholesterol was identified by comparing the sample retention time with the standard, and quantification by the corresponding peak areas was performed by external standardization, using a Sigma-Aldrich® cholesterol standard (Cholesterol, code C8667).
Statistical analysis
The obtained results were evaluated through analysis of variance and regression, using the Statistical Analysis System - SAS software (SAS, 2003). The statistical models were chosen according to the significance of the regression coefficients, using the F test at 5% probability and coefficient of determination (R2), according to the following statistical model:
Yijk= µ + li + cj + tk(ij) + eijk
Where:
Yijk= observed value of the variable;
µ= general average;
li = effect of line i;
cj = effect of column j;
tk(ij) = effect of treatment k;
eijk = random error (residual).
Results
For the concentrations of saturated fatty acids, butyric (C4:0), caproic (C6:0), caprylic (C8:0), capric (C10:0) and hendecanoic (C11:0), no significant difference (P>0.05) was observed with the inclusion of different levels of LC in the diet of confined dairy cows (Table 3).
Table 3: Saturated fatty acid composition of dairy cows fed on different levels of inclusion of licuri cake
Fatty acids1 | licuri cake levels (%DM) | Eq.2 | CV%3 | P4 | |||
0.0 | 5.5 | 11.0 | 16.5 | ||||
C4:0 | 12.6 | 12.9 | 13.6 | 12.9 | 13.0 | 9.6 | 0.7 |
C6:0 | 12.2 | 11.9 | 12.7 | 11.5 | 12.1 | 8.1 | 0.5 |
C8:0 | 8.0 | 7.7 | 8.1 | 7.2 | 7.7 | 7.3 | 0.2 |
C10:0 | 19.1 | 17.8 | 19.2 | 17.1 | 18.3 | 6.1 | 0.1 |
C11:0 | 3.1 | 2.8 | 2.9 | 2.5 | 2.8 | 11.5 | 0.1 |
C12:0 | 24.2 | 25.7 | 30.4 | 32.4 | 5 | 7.4 | 0.0 |
C13:0 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 | 13.1 | 0.6 |
C14:0 | 71.4 | 72.1 | 76.9 | 74.1 | 73.6 | 4.4 | 0.2 |
C15:0 | 8.7 | 6.9 | 6.8 | 6.1 | 7.1 | 17.7 | 0.1 |
C16:0 | 227.7 | 242.6 | 233.7 | 222.5 | 231.6 | 8.3 | 0.5 |
C17:0 | 16.1 | 15.7 | 15.3 | 13.9 | 15.3 | 10.8 | 0.4 |
C18:0 | 36.7 | 33.8 | 34.9 | 35.7 | 35.3 | 10.1 | 0.7 |
C20:0 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 8.8 | 0.9 |
C21:0 | 0.7 | 0.6 | 0.6 | 0.6 | 0.6 | 12.2 | 0.4 |
1Usual nomenclature expressed in mg g-1 fat, Butyric (C4:0), Caproic (C6:0), Caprylic (C8:0), Capric (C10:0), Hendecanoic (C11:0), Lauric (C12:0), Tridecanoic (C13:0), Myristic (C14:0), Pentadecenoic (C15:0), Palmitic (C16:0), Margaric (C17:0), Stearic (C18:0), Arachidic (C20:0), Henecosanoic (C21:0), 2Regression equations, 3Coefficient of variation, 4Probability of error and 5Y = 0.5316x + 23.804, R² = 0,96.
For lauric acid (C12:0), there was significant effect (P<0.05) with the inclusion of different levels of LC. There was at growing linear effect on the concentration of lauric acid in milk samples, resulting from the by-product (Table 2), which contributed 0.0, 1.5, 2.7 and 3.6 % among treatments.
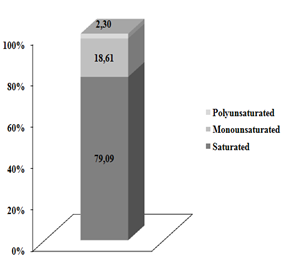
Milk saturated fatty acids represent more than 80 % in their composition, which demonstrate the biohydrogenation capacity of cattle in transforming monounsaturated and polyunsaturated fatty acids into saturated fatty acids, even with the diet contributing with more than 52 % monounsaturated and polyunsaturated fatty acids (Table 2). Among the saturated fatty acid profiles, the most abundant were palmitic acid (C16:0), myristic acid (C14:0) and stearic acid (C18:0), palmitic acid had the highest mean concentration among treatments, 231.60 mg g-1.
Table 4 shows the mono and polyunsaturated fatty acids of milk samples. There were no significant differences (P>0.05) between the inclusion levels of LC in the diet, on the fatty acid profile of myristoleic, pentadecenoic, palmitoleic, 10-heptadecenoic, oleic, linoleic, eicosatrienoic and arachidonic acids. Among these fatty acids, myristoleic (C14:1) and oleic (C18:1n9c) had higher mean concentrations, 10.2 and 78.1 mg g-1, respectively. As for the fatty acids Elaidic and Gamma-Linolenic, a linear decreasing effect (P<0.05) was observed, with a reduction of 0.1 and 0.2, respectively, for each unit (mg g-1) of LC added in the diet.
Table 4: Mono and polyunsaturated fatty acid composition of dairy cows fed on different inclusion levels of licuri cake
Fatty acids1 | Licuri cake levels (%DM) | Eq.2 | CV%3 | P4 | |||
0.0 | 5.5 | 11.0 | 16.5 | ||||
C14:1 | 10.3 | 10.4 | 10.5 | 9.7 | 10.2 | 7.4 | 0.5 |
C15:1 | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 | 25.5 | 0.9 |
C16:1 | 2.9 | 2.7 | 2.7 | 2.7 | 2.7 | 4.8 | 0.1 |
C17:1 | 1.4 | 1.2 | 1.1 | 1.1 | 1.2 | 11.5 | 0.2 |
C18:1n9t | 5.2 | 4.7 | 3.9 | 3.6 | 5 | 15.0 | 0.0 |
C18:1n9c | 90.6 | 86.0 | 51.7 | 84.2 | 78.1 | 8.8 | 0.5 |
C18:2n6 | 2.1 | 1.8 | 1.7 | 1.9 | 1.9 | 18.6 | 0.6 |
C18:3n6 | 9.4 | 8.2 | 7.0 | 5.9 | 6 | 9.2 | 0.0 |
C20:3n3 | 0.5 | 0.4 | 0.4 | 0.4 | 0.4 | 8.9 | 0.6 |
C20:4n6 | 1.1 | 1.0 | 1.0 | 1.0 | 1.0 | 8.2 | 0.3 |
1Usual nomenclature expressed in mg g-1 fat, Myristoleic (C14:1), Pentadecenoic (C15:1), Palmitoleic (C16:1), 10- Heptadecenoic (C17:1), Elaidic (C18:1n9t), Oleic (C18:1n9c), Gamma-linoleic (C18:2n6), Gamma-linolenic (C18:3n6), Eicosatrienoic (C20:3n3), Arachidonic (C20:4n6), 2Regression equations, 3Coefficient of variation, 4Probability of error. 5Y = -0.1042x + 5.197, R² = 0.97 and 6Y = -0.214x + 9,378, R² = 0,99.
The levels of CLA (C18:2cis9trans11) and (C18:2trans10cis12) in milk were influenced by the inclusion levels of LC in the diets (P<0.05) and, in the treatment with 16.5 % by-product inclusion level (Table 5), CLA levels reduced 0.01 and 0.03 mg g-1 fat, respectively.
Table 5: Conjugated linoleic acid composition and milk fat of dairy cows fed on different inclusion levels of licuri cake
Fatty acids1 | Licuri cake levels (%DM) | Eq.2 | CV%3 | P4 | |||
0.0 | 5.5 | 11.0 | 16.5 | ||||
CLA C18:2c9t11 | 0.5 | 0.5 | 0.4 | 0.3 | 5 | 7.8 | 0,0 |
CLA C18:2t10c12 | 2.1 | 1.7 | 1.6 | 1.4 | 6 | 7.8 | 0.0 |
Composition (%) | |||||||
Fat | 4.4 | 5.0 | 4.9 | 4.9 | 4.8 | 8.2 | 0.2 |
1Usual nomenclature expressed in mg g-1 fat, Conjugated linoleic acid (CLAcis9trans11) and (CLAtrans10cis12), 2Regression equations, 3Coefficient of variation and 4Probability of error. 5Y = -0.0147x + 0.534, R² = 0.99; 6Y = -0.0347x + 1.954, R² = 0,94.
There was no significant difference (P>0.05) for saturated fatty acids, monounsaturated, omega 3 and the omega 6/omega 3 ratio, as a function of the tested levels of LC (Table 6). The sum of polyunsaturated fatty acids and omega 6 decreased (P<0.05). Regarding the polyunsaturated/saturated fatty acids ratio, there was a linear decreasing effect (P<0.05). Regarding the cholesterol analysis of milk samples, there was no statistical difference (P>0.05) in cholesterol concentrations between the evaluated treatments, with a mean of 2.1 mg 100 mL-1, as can be seen in Table 6. This result followed the same effect for fat composition in milk (Table 5).
Table 6: Fatty acid sum and milk cholesterol of dairy cows fed on different inclusion levels of licuri cake
Fatty acids1 | Licuri cake levels (%DM) | Eq.2 | CV%3 | P4 | |||
0.0 | 5.5 | 11.0 | 16.5 | ||||
Saturated | 441.5 | 451.2 | 455.9 | 437.3 | 446.5 | 5.8 | 0.7 |
Monounsaturated | 111.3 | 105.9 | 100.7 | 102.3 | 105.0 | 7.4 | 0.3 |
Polyunsaturated | 15.5 | 13.5 | 12.1 | 10.9 | 5 | 8.1 | 0.0 |
PUFA / SFA8 | 0.3 | 0.2 | 0.1 | 0.0 | 6 | 9.3 | 0.0 |
n-39 | 0.5 | 0.4 | 0.4 | 0.4 | 0.4 | 8.9 | 0.6 |
n-610 | 12.5 | 10.9 | 9.7 | 8.8 | 7 | 9.8 | 0.1 |
n-6 / n-311 | 27.9 | 26.1 | 22.1 | 21.4 | 24.4 | 15.6 | 0.2 |
Cholesterol (mg 100 mL-1) | 2.3 | 2.1 | 1.9 | 2.2 | 2.1 | 14.1 | 0.4 |
1Usual nomenclature expressed in mg g-1 fat, 2Regression equations, 3Coefficient of variation, 4Probability of error, 5Y = -0.2793x + 15.309, R² = 0.99, 6Y = -0.0006x + 0,0338, R² 0.91, 7Y = -0.2278x + 12.382, R² = 0.99, 8Ratio polyunsaturated/saturated fatty acids, 9Totals omega-3: 20:3n3, 10Totals omega-6: 18:2n6, 18:3n6, 20:4n6 and 11Ration between omega-6 and omega-3.
Discussion
Fatty acid concentrations C4:0, C6:0, C8:0, C10:0 and C11:0 were not altered, probably due to the de novo synthesis and the reduced number of acetate and β-hydroxybutyrate synthesis precursors resulting from fermentation in the rumen, the main metabolic pathway involving acetyl-CoA carboxylase(13).
The increase in lauric acid concentration of LC was sufficient to alter the concentrations of the lipid profile of milk samples, from the inclusion levels used in the diet. The major contributions of lauric acid in milk may be related to the inclusion of foods from coconut(14), which have an expressive amount of (C12:0), and may be a probable justification for the results found in this study, corroborating those of(15), who evaluated palm cake for confined dairy cows. In studies conducted by(16), they reported that lauric acid is considered to be responsible for the negative effects on human health, since it is a fatty acid present in milk, mainly of animals that consumed diets containing coconut or its by-products.
Among the saturated fatty acids, the most abundant palmitic and myristic what may have been influenced by the higher percentages of these fatty acids present in LC. Palmitic and myristic acids have the potential to act negatively on human health, since they can be rapidly incorporated into cellular triglycerides, contributing to an increase in cholesterolemia(17). Stearic acid can be converted into a mono and polyunsaturated fatty acid by ruminants(18,19) Saturated fatty acids are capable of raising LDL (low-density lipoprotein) levels in human blood(20).
The absence of significance among mono and polyunsaturated fatty acids may be associated with the behavior of the Δ9-desaturase enzymatic activity in the mammary gland. According to some authors(21), Δ9-desaturase is responsible for the conversion of saturated fatty acids to monounsaturated fatty acids.
The results for fatty acids Elaidic and Alpha-Linolenic demonstrate that the profile of these acids in milk can be altered by modifications in the ruminal fermentation pattern through the action of microorganisms, and the main are Butyrovibrio fibrisolvens and Anaerovibrio lipolytica, able to hydrolyze ester bonds(22), minimizing the toxic effects of fatty acids. Even with increasing levels of by-product in the diet (Table 2), there was a reduction in (C18:1n9t), demonstrating the biohydrogenation capacity of ruminants acting as a defense mechanism.
The reduction in CLA with the inclusion of LC indicate that the milk obtained from confined animals, receiving sugarcane as a bulk source, supplemented with LC in the diet, showed that the increase in by-product partition negatively affects the concentration of conjugated linoleic acid. Trans-10 cis-12 CLA isomers are the main fatty acids responsible for the milk fat depression syndrome (SARA) but, although these isomers are present in the fat of the evaluated samples, their concentrations were low, which apparently could not act deleteriously on the de novo synthesis of fatty acids and, consequently, were not able to influence fat composition(20).
This decrease in CLA content in milk samples was more accentuated from the inclusion levels of the by-product, even though it contained 44.9 % polyunsaturated fatty acids (Table 2), they were not sufficient to increase the levels of CLA in milk. Thus, low levels of C18:2n-6 and C18:3n-6 intake in the diet contributed to a decrease in duodenal flow and, consequently, a decrease in CLA concentrations in milk. Conjugated linoleic acid is produced through the incomplete biohydrogenation of linoleic and linolenic acids in the rumen(23) CLA has shown beneficial effects on human health, which are mainly attributed to two of its isomers: cis-9, trans-11 and trans-10, cis-12(24).
The reduction of polyunsaturated fatty acids and omega-6 reflects the consumption of polyunsaturated fatty acids (Table 2) ruminants do not synthesize C18:2 and its isomers(25), especially those of the omega-6 and omega-3 families, which are obtained through the diet.
This decrease among treatments was lower than the proportion recommended by Wood et al(26), which should be above 0.4 polyunsaturated fatty acids for saturated fatty acids. Therefore, further research is necessary to improve the PUFA:SFA ratio in milk. Although the dietary balance between PUFA (omega 3) is formed from alpha-linolenic acid (C18:3) and omega 6 is formed from linoleic acid (C18:2)(27), they are considered essential for humans, in addition to conjugated linoleic acid (CLA)(9).
One of the main reasons that may explain the high concentration of saturated fatty acids in milk samples (Figure 1) is the significant reduction in PUFA consumption from LC (Table 2), as well as the biohydrogenation process. Different results were observed by others(28), in a comparative study of fatty acids of bovine and buffalo milk, which had 74.2 and 75.3 % saturated fatty acids in their composition.
Figure 1. Saturated, polyunsaturated and monounsaturated percentage fatty acids in the milk of dairy cows fed different inclusion levels of licuri cake
Conclusions and implications
The inclusion levels of LC modified the lipid profile of milk, mainly the concentrations of polyunsaturated fatty acids, conjugated linoleic acid isomers (CLAs) and Omega 6 fatty acids, which is not interesting from the human nutrition point of view. Cholesterol levels remained unchanged, regardless of inclusion level.
Acknowledgements
The authors are grateful to the Coordination for the Improvement of Higher Education Personnel (CAPES) and Program Postgraduate in Animal Science (UESB) for their support. Thanks to the Research Support Foundation of the State of Bahia (FAPESB), Pronex Project PNX0004 / 2014 and the Postgraduate Program in Animal Science (UESB) for their support.
Literature cited: