https://doi.org/10.22319/rmcp.v11i1.4740
Article
Effect of a Pennisetum purpureum and Tithonia diversifolia silage mixture on in vitro ruminal fermentation and methane emission in a RUSITEC system
Johanna Mazabel c
Steven Quintero c
Jairo Mora-Delgado b
a Universidad Nacional de Colombia - Palmira. A.A 237, Palmira, Colombia.
b Grupo de Investigación Sistemas Agroforestales Pecuarios, Universidad del Tolima, A.A. 546, Ibagué, Colombia.
c International Center for Tropical Agriculture (CIAT), A.A. 6713, Cali, Colombia.
d Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán (INCMNSZ), Departamento de Nutrición Animal, Ciudad de México, México.
*Corresponding author: vholguin@ut.edu.co
Livestock diets in the tropics are traditionally based on grass species, with significant consequent nutritional limitations. Balancing the diet requires supplementation with forage species offering high quality protein. An artificial rumen simulation technique system (RUSITEC) was used to digest a mixture of the grass Pennisetum purpureum (PP) and the sunflower Tithonia diversifolia (TD). Evaluations were done of the effects of added lactic acid bacteria on rumen kinetics and methane (CH4) release. Four treatments were analyzed: T1) a control with 100% PP silage with no inoculum; T2) a PP/TD (67:33%) mixture with no inoculum; T3) a PP/TD (67:33%) mixture inoculated with the lactic acid bacteria (LAB) Lactobacillus paracasei T735; and T4) a PP/TD (67:33%) mixture inoculated with the commercial Sil-All®4x4 LAB. Data were analyzed with a completely random design. Ammonium concentration did not differ (P>0.05) between the treatments (T2, T3 and T4) and the control (T1). The T4 treatment lowered volatile fat acids by 57 % compared to the control (P<0.001). Methane release was also lower (P<0.05) in T4 (1.36 mmol/g) than in the control (2.43 mmol/g), although levels were also relatively low in T3. The ciliate protozoa population did not differ among the treatments or with the control (P>0.05). The decrease in methane emission per gram dry matter in the PP/TD silages was probably due to lower fiber degradation levels. This suggests that methane emissions were lower per unit of edible animal protein produced even though the total amount of methane released did not differ.
Keywords: Defaunation, Fiber, Protozoa, Digestion, Silage.
Introduction
Vascular plant genetic biodiversity per unit of area is highest in the world’s tropics. Despite this wealth, animal feeding systems in tropical regions are mainly based on a very few plant species, with a limited range of forage trees and bushes(1). Worldwide demand for meat and dairy products is projected to greatly increase as human populations continue to grow. Raw materials for animal feed are a primary drive behind higher livestock production costs, highlighting the need to exploit regional biodiversity in the search for alternative feed sources and rationally employ natural local resources(2).
Livestock producers in the tropics have traditionally used diets based on associations of native and/or introduced grasses. The high cell wall content of grasses creates significant nutritional limitations that translate into low intake rates and consequent shortages in digestible nutrients intake. Both phenomena are generally due to poor microbial fermentation of grasses which results in nutrient flow and absorption lower than that required by ruminants(3). Nutritional deficiencies require the use of supplementary feeds, introduction of forage legumes in pastures, or supplementation with shrub legume forage or other non-legume high protein quality woody species(4).
Mixing a selection of forage species into the ruminant diet has been proposed as an efficient alternative for improving protein and energy supply in livestock production in the tropics(5). This can be accomplished by supplying fodder mixtures through browsing, cutting and hauling or forage conservation by silage processes(6). An appropriate energy:protein mixture for livestock is 70:30, and forage mixtures commonly include grasses and legumes in this proportion(7). Protein sources can be non-legumes from among the immense diversity of forage trees in the tropics(8). Wild sunflower Tithonia diversifolia (TD) is widely used in animal feed due to its high protein content, high digestibility and ease of management. In one study DM content in TD ranged from 13.5 to 25.0 %, depending on age and cut frequency, and crude protein values varied from 11.0 to 29.8 %, with significant differences in leaves at 30 and 60 d(9). The leaves of TD are reported to have high crude protein content (20.6 %)(10).
Silage is a nutrient conservation strategy based on anaerobic fermentation by Lactobacillus genus bacteria(11). Limited data is available on TD’s medium acidification process and ease of silage. Research is needed on the acidifying potential of native Lactobacillus strains(6), and silage stability and nutrient loss reduction once silage is exposed to aerobic conditions(12). There are reports indicating that optimization of the anaerobic fermentation process can be induced by mixtures (especially those including TD) that improve feed characteristics(6,10).
Ruminal fermentation gas production simulation techniques (e.g. continuous cultures such as the RUSITEC artificial rumen simulation system) are commonly used in in vitro experimental procedures for studying ruminal fermentation processes. They allow control of the conditions under which fermentation occurs and of the factors affecting it(13). Simulation methods are based on maintaining small quantities of ruminal fluid under controlled environmental conditions (anaerobic, temperature and kinetic) so the microbiota can act at discretion on the raw material being tested(14,15). A properly prepared simulation system can maintain a normal bacterial population in ruminal fluid under strictly controlled conditions for long time periods(16). Developed by Czerkawsky and Breckeridge(17) and modified by Machmuller et al(18), this system is specifically for quantifying fermentation by simulating the physiological activities of ruminal digestion for relatively long time periods. The RUSITEC system allows analysis of fermentation in vitro for periods sufficiently long to provide evidence of possible microbe adaptation within the rumen. The present study objective was to evaluate the association of Pennisetum purpureum (PP) and Tithonia diversifolia (TD) as a forage mixture, as well as the effect of addition of lactic acid bacteria (LAB) on ruminal kinetics and methane production variables in the RUSITEC rumen simulation system.
Material and methods
The analyses were done at the Forage Quality Laboratory of the International Center for Tropical Agriculture (Centro Internacional de Agricultura Tropical - CIAT) in the Municipality of Palmira, Columbia (3°30’09” N; 76°21’18” W).
Forages
Biomass of TD was quantified in the pre-flowering stage (60 d) for a cut made 40 cm above ground level, including leaves and stems, in February 2013. The TD was grown at the experimental farm of the National University of Colombia (Universidad Nacional de Colombia), Palmira (1,000 m asl, 24 °C average annual temperature, 1,020 mm average annual rainfall precipitation and 72 % average annual relative humidity). Collection of PP was done at 75 d of age (10 cm above ground level). Both forages were reduced to a 2-3 cm particle size with a three-blade mill (7.5 hp, 1400 rpm, and 4.5 amps; Gaitan).
Silage preparation
When ensiled, humidity in the TD was 30 % of DM and that in the PP forages was 35 % of DM. Four treatments were used: T1) Control, 100% PP silage no inoculum; T2) mixture of PP (67%) and TD (33%) silage no inoculum; T3) mixture of PP (67%) and TD (33%) silage, LAB inoculum (Lactobacillus paracasei - T735 previously isolated from TD); and T4) mixture of PP (67%) and TD (33%) silage, commercial Sil-All®4x4 inoculum [LAB mixture: Streptococcus faecium (National Collection of Microorganism Cultures – CNCM; I-3236), L. plantarum (CNCM I-3235), Pediococcus acidilactici (CNCM I-3237) and L. salivarius (CNCM I-3238)] (Table 1). The mini silos were done in triplicate. Both inocula were applied at a 104 CFU/g concentration. The forage mixtures (1,000 g each) were vacuum-packed following the Rostock model for silages(19), and stored for ninety days in darkness at room temperature (25 °C). On 90, the min silos were opened, and the silages lyophilized and ground (Thomas Wiley Mill 4, with a 1.0 mm sieve).
Table 1: Treatments analyzed in mini silo digestion process based on mixtures of Pennisetum purpureum (PP) and Tithonia diversifolia (TD)
T1= Control, 100% PP silage no inoculum; T2= mixture of PP (67%) and TD (33%) silage no inoculum; T3= TD/PP (33/67%) mixture silage, LAB (Lactobacillus paracasei - T735); and T4= TD/PP (33/67%) silage, commercial Sil-All®4x4.
In vitro ruminal fermentation in RUSITEC system
The RUSITEC system consists of eight fermenter tubes that allow simultaneous evaluation of a maximum of eight treatments. In the present study four treatments, two repetitions per treatment, were analyzed; the run effect was used as a repetition. Experimental period was ten days, of which the first four days corresponded to the adaptation period of the microorganisms to the experimental diets and the following six days were for data collection and sampling.
Ruminal fluid (890 ml) and buffer solution or artificial saliva (110 ml) were added to each of the eight fermenter tubes(20). Before being added, the ruminal liquid was filtered through four layers of gauze. Artificial saliva flow was controlled with a pump to ensure a continuous supply of 500 ml per day per fermenter, equivalent to a 0.5 per day dilution rate.
At the beginning of each experimental period, 60 g ruminal content and 16 g DM separately packed experimental silage were added to each fermentation tube for incubation. The diets were packed in nylon bags measuring 13.5 x 6.5 cm with 100 µm pores (NItex 03-100 / 32, SEFAR, Heiden, Switzerland). Subsequently, every 24 h one of the bags was replaced by a new one, starting with the ruminal content bag. In this way each bag was incubated for 48 h. After a bag change, air in the fermenter was displaced with N2 to restore anaerobic conditions. The removed bags were washed with cold water until the wastewater ran clear, and stored at -20 °C until analysis.
The fermenters were fed daily. Four hours before feeding, ruminal fluid samples were taken to quantify redox potential, pH, ammonium concentration and microorganism count. Gas collected from the fermenters was stored in 10 l bags (Supel™ Inert Foil Gas Sampling Bags, Screw Cap Valve) and quantified by water displacement.
Chemical analysis
Chemical composition analyses were run on the samples before digestion in the RUSITEC and on the incubation residues after digestion. Bromatological analyses were done following established methods: nutritional quality, NFTA Method 2.1.4 in oven at 105 °C (930.15)(21); acid detergent fiber (ADF), NFTA Method 4.1 (973.18)(22); neutral detergent fiber (NDF)(23,24,25); and crude protein (CP), Kjeldahl (984.13)(22). Estimated dry matter digestibility (EDMD) was calculated with the equation: EDMD = 88.9 - (0.779 x ADF).
Ammonium concentrations were measured following the Ammonia (NH3) Electrode Instruction Manual. Three points (1, 10 and 100 mmolar) were calibrated. For each calibration 20 ml molar solution was stirred and 1 ml sodium hydroxide added, and this repeated for the other two points (10 and 100 mmolar). Once calibrated, a 1 ml inoculum sample was taken, 18 ml distilled water and 1 ml sodium hydroxide added, and the reading taken.
pH and redox potential
Redox potential and pH were measured with a pH/ion meter (SG8, Mettler-Toledo series: B 337764747). After device calibration, a 6 ml sample was taken from each fermenter and the measurement taken with the indicated electrode.
Protozoan count
Protozoa counts were done using 2,000 µL Hayem solution (2.5 g/L HgCl2; 25 g/L Na2SO4; 5.0 g/L NaCl) and 500 µl ruminal fluid (days 0 and 1), or 100 µl Hayem solution and 1,000 µl ruminal fluid (d 2 to 8). The mixtures were placed in plastic tubes, these placed in a 0.1 mm deep Neubauer chamber (Hausser Scientific), and the protozoans present in the entire chamber area counted(26).
Methane
Methane (CH4) was measured with a chromatographer (Shimadzu GC-2014). The column (Shimadzu) had a series of columns packed for methane: 4m H-D 80/100, 0.7m S-Q and 1.5 P-N. Column temperature was 80 °C and detector temperature was FID 25 °C. This device does not employ a traditional injection port and injection is directly connected to a cable, which is at room temperature. The carrier gas was nitrogen at a column flow rate of 30.83 ml/min.
Statistical analysis
Data were analyzed using a completely randomized design (P<0.05). The variables were evaluated with general and mixed linear models for repeated measurements (Infostat, ver. 2010 software). The model included the fixed effects of treatment, measurement time and their interaction. Incubation time was considered a random effect. Different covariance structures were analyzed for each variable and the best model estimated based on Akaike (AIC) and Bayesian (BIC) information criteria.
Results and discussion
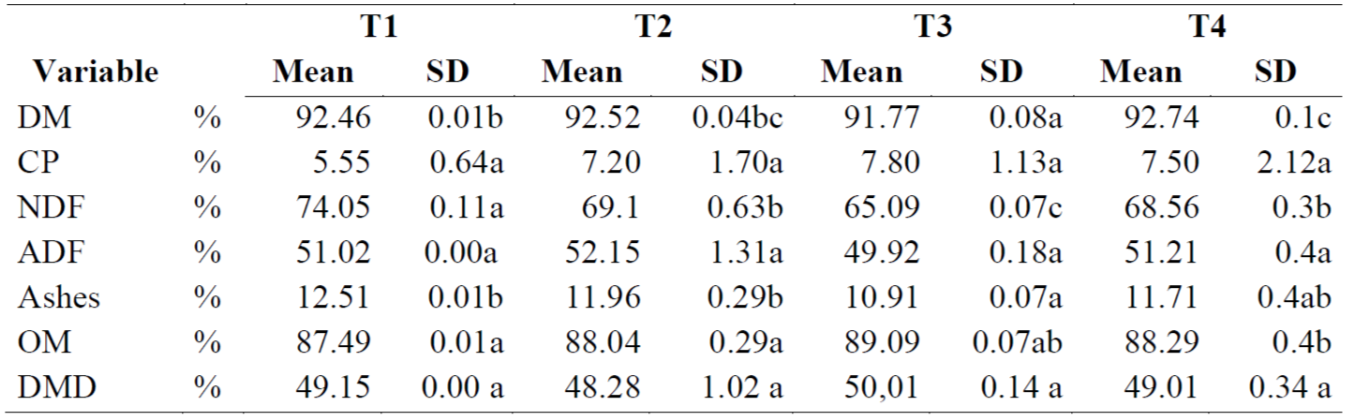
Analysis of silage chemical composition after the RUSITEC digestion process found no differences (P= 0.5182) in CP levels between silages (Table 2). Addition of TD may not have been sufficient to modify this parameter, and higher proportions of TD in the silages could have generated a significant difference between treatments. The highest CP content (7.8 %) was in T3 (PP/TD; T735), which is similar to the 6 % (30 d) and 8 % (90 d) CP reported for silage(27).
Table 2: Analysis of silage chemical composition after the RUSITEC digestion (values expressed as a percentage of dry matter)
DM= dry matter; CP= crude protein; NDF= neutral detergent fiber; ADF= acid detergent fiber; OM=organic matter; DMD=dry matter digestibility.
ab Values with different superscript are different (P<0.05).
Neutral detergent fiber (NDF) levels were highest in the control (T1; P<0.0001) and lowest in T3. Unlike CP, the fiber content in PP versus TD in the mixture significantly modified NDF, substantially lowering its levels. In the mini silo systems NDF responded inversely to TD inclusion, decreasing as TD inclusion increased. Content of NDF in forages generally ranges from 30 to 80% and the higher the fiber proportion the lower the productivity of animals that consume it(28). This dynamic is associated with DM intake since the higher the fiber content the longer the rumen retention time and the lower the voluntary fodder intake. The NDF content in treatments T2, T3 and T4 was between 15 and 19 points higher than the 54.5% reported for silages containing TD, and their ADF content was notably higher than the 32% at 90 days reported elsewhere(27). Treatment had no apparent effect on ADF, although it tended to decrease in T3. This lack of a difference in ADF may be due to the maturity of the PP, which was older (75 days) than the TD forage used in a previous report(27). Ash content was lower in T3 than in T1 and T2 (P<0.0141), and organic matter content was higher in T3 than in T1 and T2.
Silage digestibility did not differ between treatments (P= 0.1311). This is noteworthy since higher NDF content normally results in lower DM digestibility, especially when diets include woody species like TD (e.g. T2, T3 and T4, included TD leaves and stems). In addition, high cell wall content in fodder is reported to cause low digestibility and can restrict use of a fodder as animal feed(8,16, 28).
Ruminal liquid
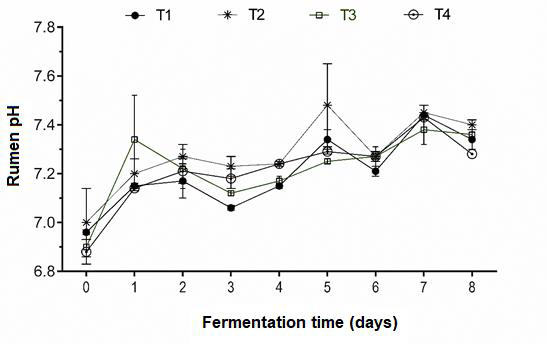
After 8 d fermentation in the RUSITEC, pH was slightly higher than normal rumen acidity range (7.3 to 7.4), but without differences (P>0.05) between treatments (Figure 1). High pH levels are likely a response to a highly fibrous diet since long fibers stimulate rumination and secretion can increase saliva production(29). Saliva functions as a lubricant for the consumed feed, and can raise pH to an average of 8.2, as well as increase sodium, potassium, bicarbonate and phosphate levels(29,30). These characteristics are simulated in the artificial saliva used in the RUSITEC, which may initially raise ruminal liquid pH, but its buffer capacity later stabilizes it. In addition to this effect of the artificial saliva, the trend from acidic towards basic pH in the present results may have resulted from a downward trend in organic acids production over time.
Figure 1. Rumen acidity in silages made from a mixture of Pennisetum purpureum (PP) and Tithonia diversifolia (TD) in a RUSITEC system
T1= Control, 100% PP silage no inoculum; T2= mixture of PP (67%) and TD (33%) silage no inoculum;
T3= TD/PP (33/67%) silage, LAB (Lactobacillus paracasei - T735); and T4= TD/PP (33/67%) silage, commercial Sil-All®4x4.
Balance in the natural rumen environment is maintained thanks to a buffer solution derived from the alkaline salivary secretion. This modulates the acidity of carbohydrate fermentation in the rumen, within a range of 5.8 to 7, generally near neutral(29). Of all the factors of the rumen medium, pH is the most susceptible to variation, and ration is the factor that can most affect it. Maintaining ruminal pH within an adequate range is the result of production and neutralization or elimination of protons in the rumen medium. Fermentation of non-structural carbohydrates (NSC) is more energy efficient but it is highly acidic; therefore the proportion of NSC in a diet needs to be limited or counteracted with structural carbohydrates (SC) which provide a buffering capacity to the rumen environment(31).
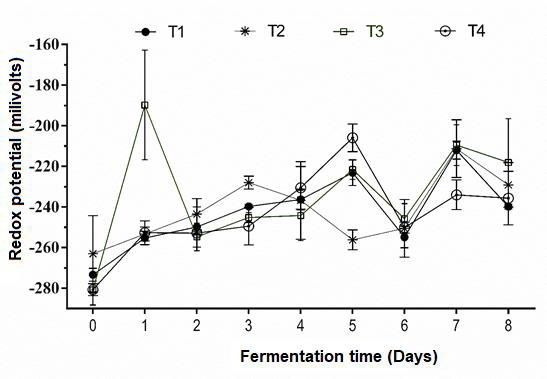
Redox potential did not differ (P>0.05) between treatments (Figure 2), which suggests that simulation in the RUSITEC closely approximated a rumen’s anaerobic environment. At times oxygen can be present in the rumen, possibly because it has entered with feed or water(3,15). Low oxygen concentrations, as indicated by negative oxidation potential (Eh) values between -250 and -450 millivolts (mV), stimulate the growth of anaerobic microorganisms(32).
Figure 2. Redox potential of silages containing Pennisetum purpureum (PP) and Tithonia diversifolia (TD) and digested in a RUSITEC
T1= Control, 100% PP silage no inoculum; T2= mixture of PP (67%) and TD (33%) silage no inoculum; T3= TD/PP (33/67%) silage, LAB (Lactobacillus paracasei - T735); and T4= TD/PP (33/67%) silage, commercial Sil-All®4x4.
Dry matter degradation
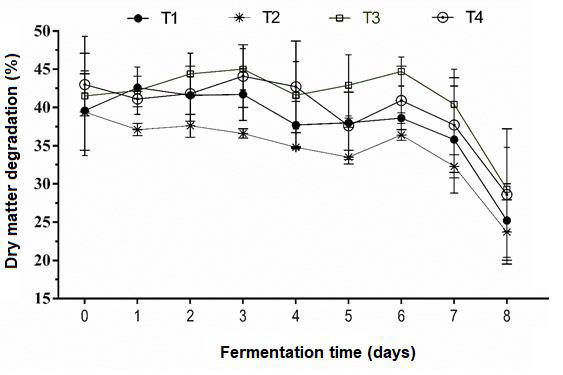
Mean DM degradation over eight days of digestion in the RUSITEC was significantly higher (P<0.0001) in T3 (41.33 ± 5.63) than in T1 (37.88 ± 5.76) and T2 (34.61 ± 4.79) (Figure 3), but did not differ significantly from T4 (39.71 ± 5.90). Longitudinal analysis also identified differences (P<0.0001) between degradation at 8 days and at the beginning of the period. The diets containing TD exhibited higher DM degradation, which can be explained by their lower non-legume protein dicotyledonous fiber content and higher forage digestibility and degradability, both inversely related to NDF content(28).
Figure 3. Dry matter degradation in silages containing a mixture of Pennisetum purpureum (PP) and Tithonia diversifolia (TD) and digested with a RUSITEC
T1= Control, 100% PP silage no inoculum; T2= mixture of PP (67%) and TD (33%) silage no inoculum; T3= TD/PP (33/67%) silage, LAB (Lactobacillus paracasei - T735); and T4= TD/PP (33/67%) silage, commercial Sil-All®4x4.
Volatile fatty acids (VFA)
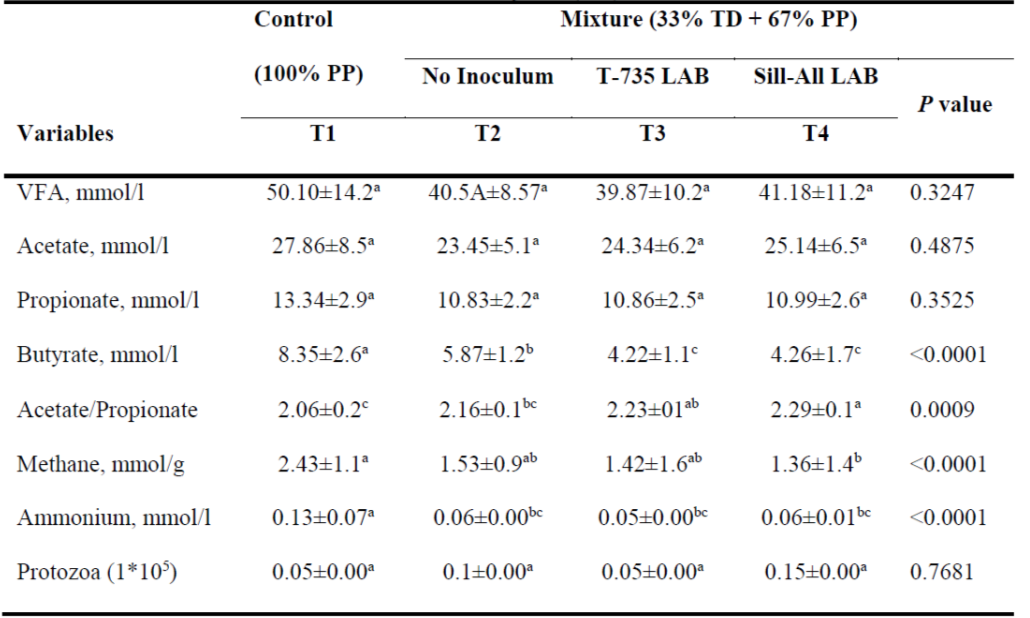
Volatile fatty acids (VFA) levels were not affected by treatment in that no differences were present, although VFA levels were slightly lower in T3 (Table 3). In forage-based diets VFA can provide between 50 and 85 % of energy requirements. Moreover, the proportions, relative to total rumen gasses, between VFA and greenhouse gases (GHG) is 65 % for CO2, 27 % for CH4 and 0.2 % for free H2(33).
Table 3: Response variables for in vitro rumen fermentation of a Pennisetum purpureum (PP) and Tithonia diversifolia (TD) mixture
T1= Control, 100% PP silage no inoculum; T2= mixture of PP (67%) and TD (33%) silage no inoculum;
T3= TD/PP (33/67%) silage, LAB (Lactobacillus paracasei - T735); and T4= TD/PP (33/67%) silage, commercial Sil-All®4x4. VFA = volatile fatty acids.
abc Different letter superscripts in the same row indicate significant difference (P< 0.05).
Fermentation product composition differed between treatments. Though not significant (P= 0.4875), acetate drainage was higher in T1 compared to the treatments containing TD. A similar trend was observed for propionate drainage, although again with no differences (P= 0.3525). Butyrate production did differ between treatments (P<0.0001), with the highest production in the control (T1), and progressively lower levels with addition of TD (T2) and the LAB inocula (T3 and T4).
In ruminants, formation of propionate is more energy efficient than that of butyrate and acetate because production of the latter two leads to release of carbon atoms that cannot be burned in the form of CH4 (or H atoms that do not convert to VFA)(34). This means that grass-only diets lead to high segregation of butyrate as C atoms are released as CH4. Total VFA was not affected by the treatments in the present study but the VFA profile was notably modified, which coincides with a previous report on diet additives in a RUSITEC(35). This is confirmed by the significant differences in the acetate/propionate ratio, which was higher in T4 and T3 (P=0.0009) than in T1. Normal VFA molar proportions in the rumen are acetic acid (65 %); propionic acid (20 %); butyric acid (13%) and others (2 %)(36). Though slightly altered, these general proportions were observed in the present results with T4 having the proportions nearest these estimates: 60.7 % (acetate); 26.7 % (propionate); 9.7 % (butyrate) and 2.9 % (others). In diets based entirely on forages the acetate:propionate ratio approaches 3:1, while as diet concentrate percentage increases the ratio narrows to 2:1 or less(16).
Ammoniacal nitrogen (NH3-N)
Production of NH3-N (as a measure of dietary N metabolism) was lower in the treatments including TD, suggesting lower dietary protein degradation in these treatments. The higher ammonium level in T1 was probably related to increased proteolysis. Daily NH3-N production changed over time in all treatments (P<0.0001); generally, values increased on the first day and then gradually decreased, which agrees with previous reports(16).
On day 8, NH3-N was highest in T1 compared to the treatments containing TD. Ammonium content did not differ among these treatments although it was slightly higher in T3, even though DM digestibility did not differ between them. This occurred despite the greater N input in T3. Previous studies have shown that in high protein forages incorporation of dietary N in microbial N increases as NH3-N decreases(37,38). This discrepancy indicates that protein degradation may occur at a slower rate in diets containing TD. Ruminal ammonia concentration in the different treatments was much lower than the 3.26 mmol/l reported elsewhere(39), but near the 1.76 mmol/l observed in an evaluation of a mixed substrate (1/3 Cratylia argentea, 2/3 Brachiaria dictyoneura) in a RUSITEC system(3). However, these levels are still lower than the 3.6 mmol/l suggested for maximum microbial protein synthesis in the rumen(40).
Methane emission
At 8-days incubation, net CH4 production per g DM was higher in T1 than in T4, which had the lowest production (Table 3). The present CH4 production was similar to the 1.47 and 1.61 mmol/g DM reported for a mixture of B. dictyoneura hay and additives(39). This also generally agrees with a study using the gas production methodology in which grass-only silages emitted high levels of CH4 which decreased as TD inclusion levels increased, and where 80 to 88 % of the total gas produced at 60 h incubation was detected at the inflection point(41). In another study of PP/TD forage mixtures CH4 production decreased as TD inclusion increased from 15 % (33.3 ml) to 30 % (30.1 ml) and 100 % (28.06 ml)(42). Decreases in CH4 emission per gram degraded MS are probably due to lower fiber degradation, suggesting that less CH4 would be emitted per unit of edible animal protein with the TD treatments, even though total CH4 emission may not decline(39).
The lower CH4 production in the PP/TD silage in the present results may be due in part to the action of condensed tannins (CT), which are known to reduce CH4 emissions in woody forage species(43). Tithonia diversifolia (TD) forage has relatively moderate CT levels(9), which can vary from 1.0 % in the dry season to 1.4 % in the rainy season(44); these are not negligible levels.
The higher CH4 production in T1 was probably due to the greater proportion of cell wall in this all-grass forage. Cell walls contain more NDF, which has low ruminal digestibility, meaning more NDF passes through the rumen into the large intestine where it can be fermented and thus produce CH4 through the action of methanogenic microbes(35,43).
Protozoa population is a good measure of rumen microbial biomass since they have relatively high weight among the microbiota. Though their net count may be smaller than the bacteria, the protozoa have a greater individual volume, resulting in a protozoan cell mass similar to the bacterial mass(16). The protozoa also colonize and degrade plant tissues in the rumen and produce enzymes capable of degrading plant and hemicellulose polysaccharides(29). In the present results the protozoa population changed over time in all treatments with values gradually decreasing during the first three days and then stabilizing during the last four days at average values that did not differ between treatments (P>0.05). Among the TD treatments (T2, T3 and T4), the protozoa count was relatively lowest in T3, which also had the highest CP proportion. This coincides with a study in which degradation was highest in substrates supplemented with a high-quality legume forage(39). However, these same forages can have high secondary metabolite (e.g. tannins and saponins) contents which can negatively affect ciliate (protozoa) populations; this would definitely improve nitrogen use but not necessarily reduce methanogenesis.
Conclusions and implications
The isolated ruminal fluid in the RUSITEC maintained variable levels comparable to the rumen in vivo. Neither the hydrogenation nor the redox potentials exhibited differences between the Control (100% Pennisetum purpureum - PP) treatment and the forage mixtures (PP/Tithonia diversifolia -TD), with and without inoculum in the silage. Values for pH were slightly higher than normal rumen acidity (7.3 and 7.4), but without variation between treatments. Redox potential did not differ between treatments. Ammoniacal nitrogen was lower in the TD treatments, which could have practical applications: reductions in the CH4 emitted per unit of animal protein produced could provide benefits even if total CH4 emissions did not decrease.
Acknowledgements
Thanks are due the Centro Internacional de Agricultura Tropical (CIAT) and the Universidad del Tolima for financial and technical support, and Patricia Ávila and Orlando Trujillo (Laboratorio de Calidad de Forraje del CIAT) for technical support.
Literature cited: